日本对虾饲料添加剂研究进展
2023-10-07张旭娟张海涛王卓铎孙广文刘敏
张旭娟,张海涛*,王卓铎,孙广文,刘敏
(1.广东恒兴饲料实业股份有限公司,广东 湛江 524000;2.农业农村部华南水产与畜禽饲料重点实验室,广东 湛江 524300)
饲料大宗原材料的短缺,使得非常规替代性原料的使用日趋增加,进而带来饲料营养不平衡、饲料含难消化碳水化合物及抗营养因子等一系列问题。减抗替抗的要求,抗生素等药物饲料添加剂的出现,对饲料和养殖业持续高质量发展提出了新的挑战。功能性饲料添加剂的使用,对解决这一系列问题尤为重要。
日本对虾(Penaeusjaponicus),也称斑节虾、花虾、车虾、花尾虾、东方褐虾等。属甲壳纲(Crustacea)、十足目(Decapoda)、游泳亚目(Natantia)、对虾科(penaeidae)、对虾属(penaeus),属甲壳尖动物,主要产于我国的渤海、黄海及朝鲜的西部沿海,是我国四大养殖对虾之一。为保持日本对虾养殖业的绿色可持续发展,选择可改善对虾生长性能、增强免疫力、降低疾病发生等的功能性饲料添加剂,至关重要。现简述了不同种类饲料添加剂在日本对虾生长、健康和品质以及饲料资源开发等方面的研究进展,以期为高效低成本功能性配合饲料的研发提供参考。
1 饲料添加剂
1.1 糖类
功能性多糖在自然界中广泛存在,具有各方面的生物活性,且安全无毒。对日本对虾的研究表明[1-3],部分免疫多糖有助于改善其免疫及生长性能。日粮中添加0.02%~2.00%的β-1,4-甘露二糖,可改善日本对虾的免疫性能,提高其对副溶血弧菌的抵抗力。但最佳剂量、投喂期和潜在作用方式,需要进一步研究。综合日本对虾质量增加率(WGR)、特定生长率(SGR)、蛋白质效率比(PER)、饲料系数(FCR)和血细胞总数、酚氧化酶(PO)活性、血清抗菌活性等指标,在0,100,500 和1 000 mg/kg 的日粮添加水平中,添加500 mg/kg 的裙带菜褐藻糖胶,效果最佳[2]。此外,日粮中单独添加0.16%的低聚糖,可显著提高日本对虾的生长性能[3](表1)。

表1 日本对虾饲料中糖类的适宜添加量
1.2 益生菌
饲料中添加益生菌,可调节水产动物体内微生态平衡,促进其生长发育,提高免疫力、抗病率。对日本对虾的研究表明,6×105cfu/g 的芽孢杆菌,可改善日本对虾的生长性能[3],而1.03×1010cfu/g 的芽孢杆菌,可改善高温环境中日本对虾的生长性能、抗氧化性能和存活率(SR)[4]。1×108~2×108cfu/g 的丁酸梭菌,可提高日本对虾肠道抗氧化性能和对高温胁迫的抵抗力,但对丙氨酸氨基转移酶和天冬氨酸氨基转移酶的活性没有显著影响[5]。1×108cfu/g 的丁酸梭菌,可提高其生长性能和对高温胁迫的耐受力,降低FCR[6];2×108cfu/g 的丁酸梭菌,诱导了α-淀粉酶基因的表达,以上2 种浓度的丁酸梭菌,均诱导了胰蛋白酶基因的表达,提高了肠道短链脂肪酸(丙酸、丁酸)和粗蛋白的含量[5]。1 g/kg 的热灭活植物乳杆菌,对日本对虾的抗应激性能改善效果显著,但0~1 g/kg 的水平,对蛋白酶活性没有显著影响[7]。
此外,维生素C+低聚糖+芽孢杆菌,显著提高了日本对虾的生长性能,提高了肌肉超氧化物歧化酶活性(SOD)、总抗氧化能力(T-AOC)、酸性磷酸酶(ACP)、碱性磷酸酶(AKP)、PO 活力以及肝胰腺中SOD、T-AOC、PO 活力[3]。地衣芽孢杆菌+枯草芽孢杆菌+异麦芽低聚糖,改善了日本对虾的肠道菌群结构和非特异性免疫性能[8](表2)。

表2 日本对虾饲料中益生菌的适宜添加量
1.3 虾青素及其他类胡萝卜素
虾青素具有增强免疫力、提高抗氧化性能、着色等多种生物学功能及作用,因而在水产养殖中应用广泛。日粮中单独添加不同水平的虾青素,在日本对虾不同生长阶段的生长性能、存活率和抗应激性能等方面的改善效果各有千秋[9](表3)。日粮中虾青素和维生素E,对日本对虾的生长性能和色素沉积具有协同作用[10];日粮中虾青素和胆固醇在存活率、生长性能、体近似成分、免疫反应和淡水应激试验方面,不存在显著的互作,但在虾体色参数和全虾的虾青素含量方面存在互作[10]。无论胆固醇是哪种水平,0.6%虾青素添加组幼虾不同部位(全身、肌肉、甲壳和头部)的色素沉积性能更好,虾青素含量更高,幼虾肝胰腺蛋白酶、脂肪酶和淀粉酶活性更高[10]。日粮中分别添加50 或100 mg/kg的雨生红球藻(含虾青素)、太平洋螺旋藻(含非虾青素类胡萝卜素)、合成虾青素,可显著提高日本对虾幼虾的存活率以及对低溶解氧胁迫的抵抗力[11]。
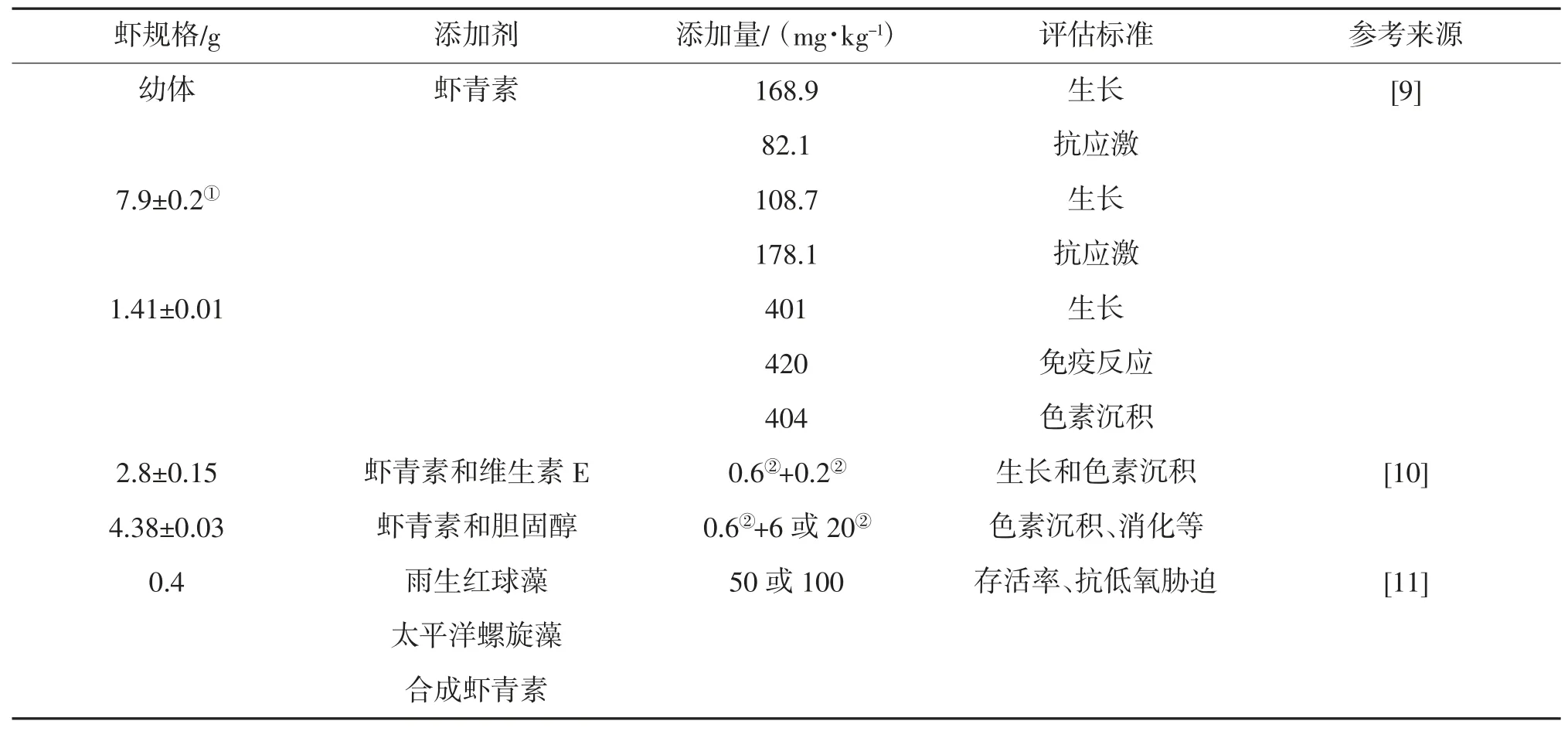
表3 日本对虾饲料中虾青素的适宜添加量
Abrill 等[12]研究了饲料添加不同比例的类胡萝卜素色素[加丽素红(0.1,0.5,2.5 和10 g/kg)和角黄素(0.01,0.05,0.25 和1.25 g/kg)],对日本对虾([10.3±3.5)g]生长性能的影响。对于肝胰腺来说,10 g/kg 加丽素红和0.01 g/kg 角黄素日粮组,类胡萝卜素的累积量最大;对于虾壳,0.5 g/kg 加丽素红和0.05 g/kg 角黄素日粮组类胡萝卜素的累积量最大。
1.4 氯化胆碱
相关研究表明,饲料单独添加氯化胆碱,在日本对虾仔虾磷脂酰胆碱和蛋氨酸等的含量[13-14],以及生长、存活和耐应激方面改善效果显著[14];而氯化胆碱与蛋氨酸、大豆磷脂酰胆碱、肌醇对日本对虾不同生长阶段的生长性能、饲料利用、体成分等方面的影响,也分别存在显著的互作(表4)。氯化胆碱和蛋氨酸在日本对虾幼虾的SR、WGR、SGR、FER、PER 以及全虾胆碱和蛋氨酸含量方面存在显著的互作。缺乏蛋氨酸的日粮中添加0.12%的氯化胆碱,可以为日本对虾幼虾补充其所需的甲基。作为甲基来源,日本对虾对胆碱的需求高于蛋氨酸[13-14]。对于仔虾,氯化胆碱和大豆磷脂酰胆碱在日本对虾的SR、全虾水分和蛋白质含量方面不存在互作,但在WG、表观FER、表观PER 和全虾总脂、游离胆碱和磷脂酰胆碱含量方面存在显著的互作。对于幼虾,两者在虾体近似成分和脂肪酸含量方面不存在互作,但在WG、SGR、SR、FER、PER、虾体游离胆碱和磷脂酰胆碱含量方面,均表现出显著的互作。胆碱是满足日本对虾仔虾和幼虾生长和存活所必需[15]。氯化胆碱和肌醇在日本对虾幼虾的SR、WGR、SGR、FER、PER 以及全虾磷脂和磷脂酰肌醇含量方面存在显著的互作。日粮中缺乏肌醇时,需添加0.12%的氯化胆碱,帮助幼虾补偿其所需的磷脂;日粮中缺乏氯化胆碱时,需要添加肌醇以补偿磷脂缺乏[16]。

表4 日本对虾饲料中氯化胆碱的适宜添加量
1.5 其他
适量的结晶氨基酸或植酸酶或鱼可溶性蛋白,均能提高日本对虾日粮中高水平植物蛋白的利用率,三者组合添加效果更佳[17](表5)。在0.25%、0.50%、1.00%的谷氨酰胺二肽添加水平中,各添加组显著提高了日本对虾的PO、ACP、溶菌酶(LSZ)活性,显著降低了肝胰腺细胞凋亡率;0.5%~1.0%添加水平,显著提高了SOD 和AKP 活性;1.0%添加水平,显著提高了总蛋白、肠绒毛高度[18]。富含β胡萝卜素和不饱和脂肪酸的海洋红酵母,可提高幼虾的成活率、生长性能和抗逆性能[19]。日粮中添加2%和10%的蜗牛壳粉,显著提高了日本对虾的抗应激能力,蜗牛壳粉添加量≤2%的日粮组,条件指数和肝体比显著更低[20]。从多毛类动物中提取的可溶性三氯乙酸、中性脂质和极性脂质组分,以0.5%(干物质基础)添加到基础日粮中,中性脂质对日本对虾亲虾的卵巢成熟最有效[21]。日粮中添加金针菇提取物麦角硫,对日本对虾的免疫系统无不利影响,但显著抑制了对虾血淋巴中的多酚氧化酶活性,降低了血细胞中酚氧化酶原基因的表达水平。因此,摄食添加了麦角硫因日粮的虾在冰冻期间,其黑变病的发展被显著抑制。虾在整个冰冻储藏期间,其提取物添加组的脂质氧化也得到了有效控制[22]。富含核苷酸的面包酵母提取物,对日本对虾具有免疫刺激效果[23]。对于缺乏蛋氨酸和赖氨酸的日本对虾幼虾,大豆分离蛋白基础日粮添加包膜蛋氨酸和赖氨酸,可有效改善这类日粮的营养价值[24]。
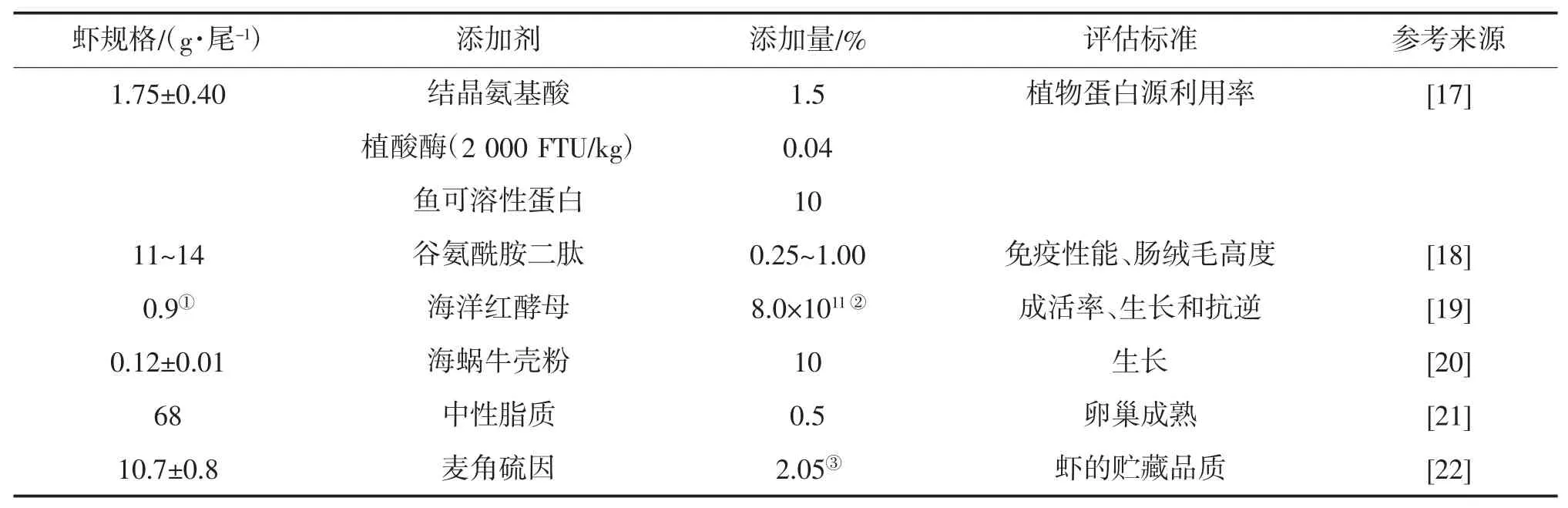
表5 日本对虾饲料中其他添加剂的适宜添加量
2 日本对虾饲料资源的开发
日本对虾属肉食性虾类,对蛋白质的需求较其他虾类高,在保障其正常生长的前提下,选择合适的饵料,以减少对水体的污染,对提升养成率非常必要。有研究表明[25],丰年虫的氨基酸组成,与对虾的氨基酸需求最为相近,相比于人工配合饲料、虾片和牡蛎肉,感染WSSV 后,摄食丰年虫无节幼体和鱼粉的日粮组,日本对虾的存活率最高。卤虫无节幼体、菲律宾蛤仔、人工配合饲料,均是常用的对虾养殖饵料。庞智予等[26]的研究表明,投喂卤虫无节幼体,日本对虾[(22.36±2.48)g]的存活率最高;投喂配合饲料,对虾生长速度最快,呈味氨基酸含量最高。相比于未富集高度不饱和脂肪酸的卤虫无节幼体,富集了HUFA 的卤虫无节幼体,显著提高了日本对虾仔虾PL4[(1.14±0.31)mg]的生长速度、同化和总转化效率[27]。韩永望等[28]比较了强壮藻钩虾和人工配合饲料对日本对虾[(2.07±0.184)g]生长和抗病力的影响,相比于人工配合饲料,钩虾可提高日本对虾的生长速度、免疫性能和抗病力;而且钩虾适应性比较强,在养殖池中繁殖速度快,能够形成对虾稳定的天然饵料。
3 结语
多糖、益生菌、虾青素、氯化胆碱等添加剂,应用于日本对虾饲料中,功效显著。但由于大多饲料添加剂的研发,均在特定试验条件下进行,而应用到养殖端,则常常达不到相应的效果。为保障日本对虾饲料与养殖业健康、绿色、高质量发展,应建立相应的标准与规范,使应用于对虾饲料添加剂产品的开发,遵循合规、安全有效、提高生产效益和改善品质等原则。
