鱼腥草bZIP基因的鉴定与分析
2023-07-06肖敬忠余小丽姚彩凤曾安杰李芙蓉付鸿博
蔡 伟,肖敬忠,余小丽,姚彩凤,陈 蔓,曾安杰,李芙蓉,陈 辽,刘 雷*,付鸿博
鱼腥草基因的鉴定与分析
蔡 伟1,肖敬忠3,余小丽3,姚彩凤3,陈 蔓3,曾安杰3,李芙蓉3,陈 辽3,刘 雷3*,付鸿博2*
1. 绵阳师范学院资源环境工程学院,四川 绵阳 621000 2. 红河学院生物科学与农学学院,云南 蒙自 661199 3. 绵阳师范学院生命科学与技术学院,四川 绵阳 621000
筛选使鱼腥草具有不同环境适应性的候选基因,为后续繁育出具有更强环境适应性的鱼腥草品种奠定基础。以鱼腥草转录组数据为基础,鉴定了基因家族成员,并对其理化性质、系统进化树、motif基序及在2个不同生态型鱼腥草中的表达进行分析。共鉴定得到163个基因,编码蛋白质的氨基酸数量在115(HcbZIP48和HcbZIP149)~703 aa(HcbZIP97),超过53%的家族成员为碱性蛋白,超过84%的家族成员为不稳定蛋白,所有家族成员均为亲水性蛋白。二级结构中,161个家族成员是由α-螺旋、延伸链、β-折叠和无规则卷曲4部分构成,β-折叠所占比例最低,α-螺旋和无规则卷曲所占比例较高。系统进化树分析将163个家族成员分成了9个类别,包括C、S、G、A、I、U、H、F和D,其中被归到D类别中的基因数量最多共有35个,数量最少的为C类别仅有4个,没有基因被归类到E和B类别中。Motif基序分析发现,motif1(RLLQNRESARRSRLRKKAYVQELESSVAK)高度保守在每个家族成员中均鉴定到且构成了基因的保守结构域。不同类别下的基因其motif基序构成较为接近,且部分motif基序只存在特定类别的基因中。表达分析发现,在鱼腥草7号药材中表达的基因数量多于鱼腥草6号药材,同时也发现单独在7号药材中表达的基因数量也多于鱼腥草6号药材。共发现6个基因,、、、、和在6号和7号药材中均呈现出较高的表达量。通过比较6号和7号的差异基因发现,共有96个差异表达基因,并将其分成了2大类,共63个差异基因在7号药材中表达量较高,33个差异基因在6号药材中表达量较高。共筛选出4个基因、、和作为鱼腥草6号和7号2种材料能够形成适应不同生态条件的候选基因。
鱼腥草;生物信息学;差异表达基因;不同生态型;基因家族
植物在其生命周期经常面临不利的环境条件,如干旱,紫外线,极端温度等。为了在这些具有挑战性的环境中生存和茁壮成长,植物经过多年的进化和适应,已经发展出各种各样的防御机制。深入了解这些机制将有助于育种者开发优良品种,供生产实践采用以更有效地应对气候变化。转录因子(transcription factors)已被证明通过自我调节和调节下游靶基因表达在植物生长和发育中起关键作用[1],并通过结合靶基因的启动子参与主要成分来响应不同的胁迫[2]。基因是植物中最大的转录因子家族之一,其家族成员已经在植物、动物和许多其他物种中被鉴定出来。含有大约70个氨基酸的bZIP结构域具有碱性区域(N-x7-R/K基序)和亮氨酸拉链区域[3]。转录因子主要分布在核蛋白植物中。从烟草中分离到基因,构建了NbbZIP28-GFP融合蛋白[4],前体蛋白从内质网膜分离并进入细胞核发挥作用[4]。转录因子家族主要识别ACGT的顺式作用元件,例如ABRE、G-box、A-box和C-box作为启动子区域[5]。随着基因的克隆和功能的发现,由于其在非生物胁迫下具有重要的调节功能,受到了研究者的广泛关注。在白桦的研究中,通过过表达后发现转基因株系的耐盐能力提升[6]。在用不同浓度PEG-6000处理糜子幼苗后,、、和这4个基因都表达量随处理浓度的变化呈现显著上调或下调,说明了转录因子参与了糜子干旱胁迫[7]。在玉米的研究中,随着NaCl处理时间的延长和的表达量显著升高,在低温(4 ℃)处理时,的表达量随处理时间的延长显著升高,说明了转录因子参与玉米盐和低温胁迫[8]。在小麦的研究中,过表达可提高植株的耐热性[9],过表达可提高植株的耐盐性[10]。综上,转录因子在植物应对非生物胁迫,包括干旱、高盐、低温和高温等方面具有重要作用。
鱼腥草为蕺菜Thunb.干燥地上部分,是一种辛辣的,心形叶状的多年生草本植物,具有很高的食用和药用价值[11]。幼嫩的茎叶是桌上的一种特殊的蔬菜[12]。鱼腥草地上部、地下部和地上部干花期的新鲜植物作为常见的药用植物[13],具有清热解毒、祛痰止咳和镇痛等功效[14-15],同时鱼腥草还是许多药物的原料,可加工成西药注射剂或称为“中药抗生素”的中成药[16],在抗菌、抗病毒方面具有良好的作用,是连花清瘟胶囊、抗病毒合剂的重要成分[17]。鱼腥草由于其较强的环境适应性,广泛分布于我国南方各省区,西北、华北地区及西藏等地区[18],但由于各地区的生长环境差异较大,各地区的鱼腥草无论是在外观还是营养成分上都会存在着一定的差异,同样也会存在不同的生态适应性[19]。本研究以2种不同生态类型的鱼腥草(6号和7号)作为材料,以转录组数据为基础挖掘鱼腥草基因家族成员,以期筛选出使鱼腥草6号和7号药材具有不同环境适应性的候选基因,为后续繁育出具有更强环境适应性的鱼腥草品种奠定基础。
1 材料
共选取2种不同生态类型的鱼腥草(6号和7号)作为实验材料。2020年9月在雅安雨城区濆江南岸的老板山脚下收集到6号药材。2020年9月在四川省温江市惠和村收集到7号药材。经绵阳师范学院生命科学与技术学院罗明华教授鉴定为三白草科植物蕺菜Thunb.,样本材料保存于该校生命科学与技术学院教学农场,在相同的场地条件和水肥管理2年后,采集地上部分(茎和叶)储存在−80 ℃冰箱用于进一步分析。
2 方法
2.1 转录组测序及差异表达基因分析
转录组测序由武汉迈特维尔生物科技有限公司完成,对于转录组测序结果,使用DESeq2方法对6号和7号2份材料进行差异表达分析,经过差异分析后,需要用Benjamini-Hochberg方法修正多重假设检验的假设检验概率(值),以获得错误发现率(FDR)。差异表达基因的筛选条件为|log2Fold Change|≥1,FDR<0.05。
2.2 鱼腥草bZIP基因的鉴定
以转录组测序结果预测的蛋白序列为基础进行鱼腥草基因的鉴定,bZIP结构域序列(PF00170)下载于Pfam数据库,以此来查询序列以比对蛋白,通过hmmer3.0软件来鉴定鱼腥草基因并结合转录组使用iTAK软件所预测出来的基因为候选鱼腥草基因。利用NCBI-CDD数据库对所有候选基因的结构域进行验证,最终确定鱼腥草基因家族成员。
2.3 鱼腥草bZIP基因的生物信息学分析
利用所鉴定的基因和已知拟南芥基因进行系统树。拟南芥数据下载于TAIR数据库,利用MEGA软件的Neighbor- Joining方法(bootstrap replications=1000,Model=p-distance)构建系统进化树。鱼腥草基因的理化性质和二级结构利用ProtParam和SOMPA在线数据库进行分析。家族成员的保守结构域利用NCBI进行分析,利用TBtools软件的Simple MEME Wrapper工具分析家族成员的motif基序,并利用TBtools软件进行可视化。
2.4 鱼腥草bZIP基因的表达分析
根据转录组数据绘制所鉴定得到的鱼腥草表达量热图和Venn图,并利用TBtools软件进行可视化。将6号和7号2个材料中差异表达的基因进行K-means聚类分析,并利用迈维在线云平台(https://cloud.metware.cn)实现可视化。
3 结果与分析
3.1 鱼腥草bZIP基因家族成员进化树分析
将所鉴定得到的163个鱼腥草基因和拟南芥进行进化树分析,并根据拟南芥的分类方式进行分类,结果表明,拟南芥基因共分成11个类别,包括C、S、G、A、I、U、E、H、F、B和D(图1)。鱼腥草基因被归类到这11个类别中的9个,包括C、S、G、A、I、U、H、F和D,其中被归到D类别中的基因数量最多共有35个,其次为A和I类别,分别有32和31个,被归类到C、S、H和U类别中的基因数量均少于10个,数量最少的为C类别仅有4个,同时可以发现,没有基因被归类到E和B类别中。

图1 鱼腥草和拟南芥bZIP基因的进化树分析
3.2 鱼腥草bZIP基因家族成员的理化性质分析
对所鉴定的163个鱼腥草基因的理化性质和二级结构进行分析(表1),编码蛋白质的氨基酸数量在115 aa(HcbZIP48和HcbZIP149)~703 aa(HcbZIP97),其中类别A中的32个蛋白的氨基酸数量均在400 aa左右,类别S中的7个蛋白的氨基酸数量较少,均在110~160 aa。在这163个HcbZIP蛋白中,相对分子质量在13 090(HcbZIP48和HcbZIP149)~75 470(HcbZIP97)。等电点在4.60(HcbZIP159)~9.96(HcbZIP136),其中有76个蛋白的等电点小于7为酸性蛋白,有87个蛋白的等电点大于7为碱性蛋白。不稳定系数在28.24(HcbZIP15、HcbZIP23和HcbZIP151)~79.99(HcbZIP153),其中有25个蛋白的不稳定系数小于40为稳定蛋白,有138个蛋白的不稳定系数大于40为不稳定蛋白。脂肪族氨基酸指数在43.37(HcbZIP17)~105.32(HcbZIP08)。亲水性系数在−1.284(HcbZIP30)~−0.065(HcbZIP79),表明所有的蛋白均为亲水性蛋白。对二级结构预测后发现,共有161个HcbZIP家族成员由α-螺旋、延伸链、β-折叠和无规则卷曲4部分构成,HcbZIP30由α-螺旋、β-折叠和无规则卷曲3部分构成,延伸链的占比为0,HcbZIP30由α-螺旋、延伸链和无规则卷曲3部分构成,β-折叠的占比为0。β-折叠在所有蛋白中的占比均较低,均低于10%,延伸链在所有蛋白中的占比均低于20%,α-螺旋的占比在15.67%(HcbZIP17)~84.17%(HcbZIP08),无规则卷曲的占比在9.57%(HcbZIP48)~75.00%(HcbZIP17)。

表1 鱼腥草bZIP基因家族成员的理化性质分析

续表1

续表1
3.3 鱼腥草bZIP基因家族成员保守结构域及motif基序分析
将所鉴定得到的HcbZIP蛋白进行单独进化树构建(图2-a),同样分成了9个类别。通过分析基因的保守结构域发现,所鉴定的163个基因均有bZIP家族的保守结构域(图3-c),进一步证实这些基因均属于bZIP家族成员。为了分析bZIP家族成员的蛋白特征,对其motif基序及组成进行分析(图2-b和图3),共设定了20个motif基序,其中motif1在163个家族成员中均被鉴定到为最保守的motif基序,它也构成了家族成员的保守结构域,motif1(RLLQNRESARRSRLR- KKAYVQELESSVAK)由29个氨基酸位点组成。在类别C和U中的基因其motif基序结构简单,只鉴定到了motif1。类别S中的基因motif基序结构也较为简单,只包含motif1和motif11。类别A中的基因除了外的其他基因的motif基序组成均较为丰富,包含了8~9个motif基序,其中motif6、motif7、motif9和motif20只在类别A中的基因中鉴定到。Motif18(HIRHQRTSSES- FIFDEQPSWLNDLLDEPETPVGRGTHRRSSSDSV-AYLDI)和motif19(RRPAIKDWETIVDTCGY- QNSVASAREWLVEKHEADRSDMHAYGY)只在类别I的部分基因中被鉴定到。Motif5(MADTSPRTDTSTDGDLDDKNQRLDPGHSVM- AVASDSSDRSK)和motif12(FDKALGSFDTASAA- RILSQTQSLNQGQESNIPVLNSNFEKW)只在类别D和F的部分基因中被鉴定到。Motif8(ILVSHLEP- LTEQQLMGICNLQQSSQQAEDALSQGMEALQQS-LAETLASGS)只在类别D的部分基因中被鉴定到。
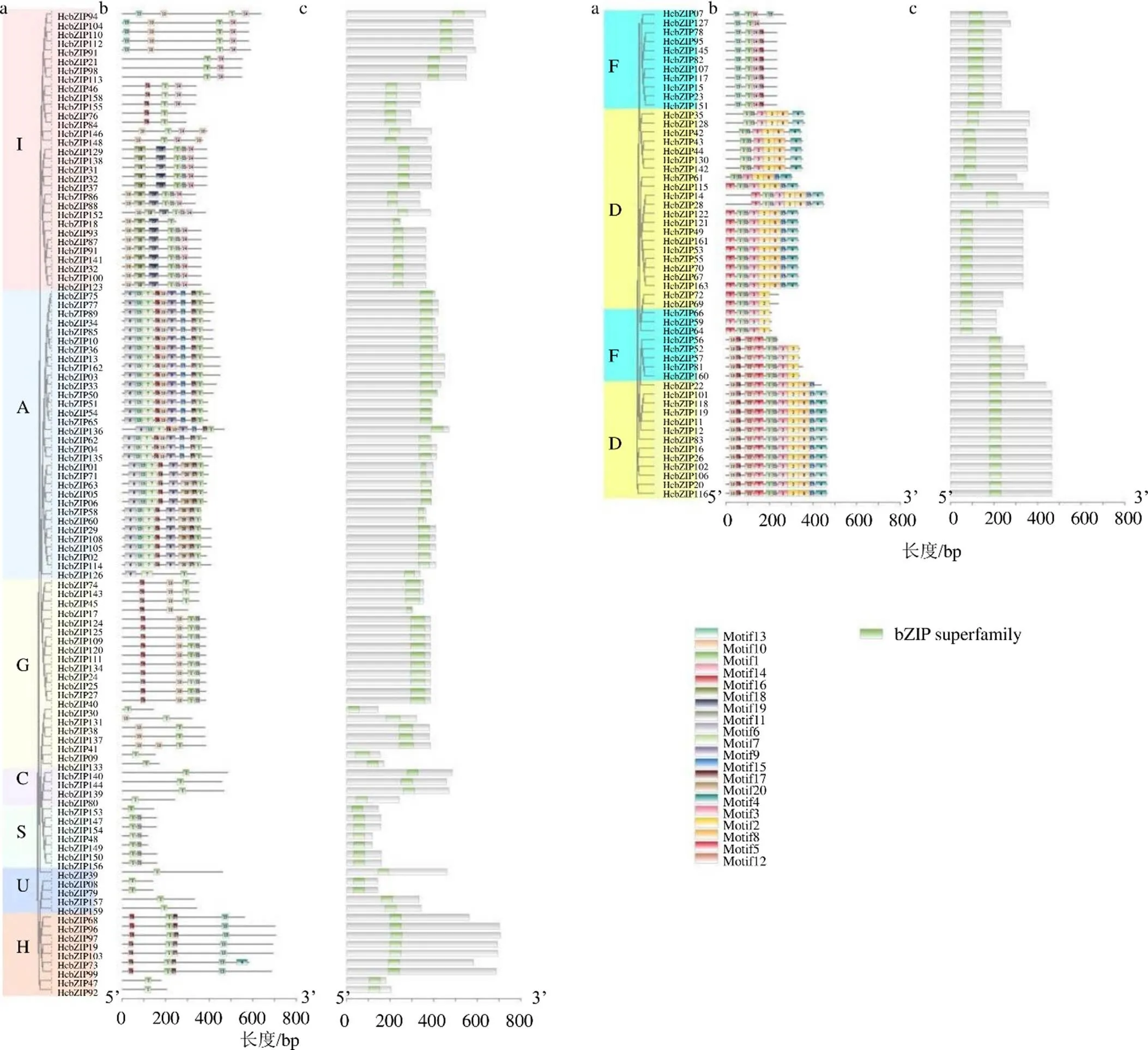
图2 鱼腥草bZIP蛋白进化树(a),motif基序(b)和保守结构域(c) 分析

图3 鱼腥草bZIP motif基序组成分析
3.4 鱼腥草bZIP基因表达分析
根据转录组数据将所鉴定得到的163个基因在6号和7号药材中的表达模式进行分析并绘制了表达量热图和Venn图(图4-A、B)。通过表达量热图(图4-A)可以发现,基因在6号和7号鱼腥草的中表现出不同的表达模式,在6号药材中表达的基因共有142个,在7号药材中表达的基因共有158个。共发现6个基因,包括、、、、和在6号和7号药材中均呈现出较高的表达量,其中和在6号药材中的FPKM值均超过100,和在7号药材中的FPKM值超过100,在6号和7号药材中所有的基因中表达量最高的均为。但大部分的基因的表达量较低,6号药材中有41个基因的FPKM值小于1,7号药材中有23个基因的FPKM值小于1。同时还发现1个基因在6号和7号药材中均不表达。通过Venn图(图4-B)可以发现,在6号和7号中均表达的基因共有138个,只在7号药材中表达的基因有20个,其中和的表达量相对较高,FPKM值大于10。只在6号药材中表达的基因有4个且表达量均较低,其中有3个基因的FPKM值小于1。

图4 鱼腥草bZIP基因在6号和7号药材中的表达模式分析(A) 和Venn图(B)
3.5 不同生态品种中bZIP基因差异表达分析
为了进一步比较在不同生态型鱼腥草材料中的表达差异,对6号和7号药材中差异表达基因进行了分析,共有96个基因在6号和7号2个品种中呈现出差异表达。通过K-means聚类分析后可以发现(图5),这96个基因被分成了2个类别,其中A类别中包括了63个差异基因,这63个基因表现出在7号药材中的表达量与6号相比为上调,其中有多个基因是在6号药材中不表达而与7号药材形成差异且差异倍数较大,在2个材料中均表达而形成较大差异倍数的基因包括和。B类别中包括了33个差异基因,这33个基因表现出在7号药材中的表达量与6号相比下调,其中是在7号药材中不表达而与6号药材形成的差异,有2个基因包括和在6号和7号药材中均表达,但形成了较大的差异倍数。

图5 鱼腥草6号和7号药材中差异表达的bZIP基因分析
4 讨论
转录因子是植物中最大和最多样化的家族之一[20]。目前,基因家族已在许多物种中被鉴定,在西瓜[21]和文冠果[20]中分别鉴定得到62和64个基因家族成员,分析后发现这2个物种中均有14对节段复制事件。在葡萄的研究中发现[22],共鉴定到55个基因家族成员,同样存在14对节段复制事件。在苹果[23]和大豆[24]的研究中分别鉴定到112和160个基因家族成员,在这2个物种中发现了更多的节段复制和串联复制事件。这些也表明节段复制和串联复制均对基因家族成员的扩增具有很大的贡献。在本研究中,共鉴定得到163个基因家族成员,由于鱼腥草基因组数据测序还未完成,本研究是基于转录组数据进行的家族成员鉴定,因此无法直接判断基因家族成员扩增的具体原因,通过以上前人的结果推测鱼腥草基因同样存在着数量较多的节段和串联复制事件。鱼腥草在长时间的不断进化过程中需要适应环境变化带来的生存压力,由于对非生物胁迫具有重要的作用,需要存储环境信息,因此可能也导致了基因的产生、复制和进化。
为了进一步研究鱼腥草基因的进化和起源,用拟南芥和鱼腥草基因进行了系统发生树分析,结果显示163个HcbZIP蛋白分为9个类别,包括C、S、G、A、I、U、H、F和D,每个类别都包含一个或多个基因。该分类类似于拟南芥[25],文冠果[20]和柳枝稷[26]。A类别的大多数功能信息提示在脱落酸或应激信号中的作用。因此,A类别的s在种子和营养组织中似乎都是ABA信号转导的重要参与者[25]。本研究共有32个基因被分到类别A中,其中在2个材料中的表达量较高且差异较大,推着该基因可能参与到鱼腥草ABA信号转导途径。S类别是拟南芥最大的类别之一,在胁迫处理后也被转录激活或在花的特定部分特异表达[25]。拟南芥家族中的S类别的基因在响应低温和干旱有重要作用[27],本研究中共有7个基因被分到S类别下,其中与6号药材相比在7号中下调表达,、、和在7号中上调表达,推测这些基因可能影响6号和7号鱼腥草对低温和干旱的响应差异。
在植物发育和应对不同生存环境方面起着重要作用。例如,几个水稻基因如被鉴定为对冷胁迫反应的调节因子[28]。西瓜的和可能在保护植物免受线虫感染和低温胁迫方面发挥重要作用[21]。文冠果中的有多个基因对盐胁迫和冻害都有反应,而和对脱落酸和低温都有反应[20]。在马铃薯中和在不同环境中均有高表达且稳定表达[29]。有研究发现,几个基因在冷处理后上调或下调表达[30]。本研究选用了原产地具有不同生态类型的鱼腥草6号和7号药材,6号和7号鱼腥草在表观形态上具有明显的差异,同时通过我们前期测定的相关生理数据发现,它们在多种代谢物含量具有明显差异,包括黄酮类物质,生物碱类物质,挥发性代谢物,这些物质上的差异均导致了这2个药材具有明显的差异。通过对基因在6号和7号鱼腥草中的表达情况进行分析,所鉴定到的163个基因中有142个在6号药材中表达,有158个在7号药材中表达,同时还发现,只在7号药材中表达的基因有20个,只在6号药材中表达的基因有4个。为了比较2个品种中表达差异,筛选得到96个差异表达基因,7号与6号相比,有63个基因在7号中表达上调,33个表达下调。结合表观形态、生态条件、物质含量及表达量的差异筛选出、、和这4个基因将作为6号和7号鱼腥草能够适应低温和干旱等不良环境的候选基因,为研究鱼腥草抗非生物胁迫的分子基础及相关抗性品种的选育提供理论数据
利益冲突 所有作者均声明不存在利益冲突
[1] Wang Z, Tang J, Hu R,. Genome-wide analysis of the R2R3-MYB transcription factor genes in Chinese cabbage (ssp.) reveals their stress and hormone responsive patterns [J]., 2015, 16(1): 17.
[2]Lindemose S, O'Shea C, Jensen M K,. Structure, function and networks of transcription factors involved in abiotic stress responses [J]., 2013, 14(3): 5842-5878.
[3]Nijhawan A, Jain M, Tyagi A K,. Genomic survey and gene expression analysis of the basic leucine zipper transcription factor family in rice [J]., 2008, 146(2): 333-350.
[4]Shen L L, Li F F, Dong W F,.NbbZIP28, a possible regulator of unfolded protein response, plays a negative role in viral infection [J]., 2017, 149(4): 831-843.
[5]Foster R, Izawa T, Chua N H. Plant bZIP proteins gather at ACGT elements [J]., 1994, 8(2): 192-200.
[6]郭依萍, 石晶静, 周美琪, 等. 白桦BpbZIP1基因抗旱耐盐分析及ABRE元件结合鉴定 [J]. 林业科学研究, 2020, 33(5): 68-76.
[7]王媚, 刘天鹏, 何继红, 等. 糜子bZIP基因家族鉴定及幼苗期聚乙二醇6000处理下的表达特征 [J]. 应用与环境生物学报, 2022, 28(4): 920-930.
[8]贾利强, 赵秋芳, 陈曙. 11个玉米bZIP基因应答逆境胁迫的表达分析 [J]. 安徽农业大学学报, 2022, 49(2): 197-204.
[9]Geng X L, Zang X S, Li H R,. Unconventional splicing of wheat TabZIP60 confers heat tolerance in transgenic[J]., 2018, 274: 252-260.
[10]Bi C X, Yu Y H, Dong C H,. The bZIP transcription factor TabZIP15 improves salt stress tolerance in wheat [J]., 2021, 19(2): 209-211.
[11]尹明星, 陈婧, 施春阳, 等. 从马兜铃内酰胺探讨鱼腥草安全性 [J]. 中草药, 2021, 52(19): 6045-6051.
[12]Liu X C, Tian J, Pan Y Z,. Structural characterization and biological activity of polysaccharides from stems of[J]., 2022, 11(22): 3622.
[13]Luo Q, Meng P H, Jiang D W,. Comprehensive assessment ofthunb., an important medicinal plant and vegetable [J]., 2022, 12(10): 2582.
[14]陈龙, 彭程琪, 樊明旭, 等. 鱼腥草药理作用及抗肺炎作用研究进展 [J]. 人参研究, 2022, 34(5): 52-54.
[15]武营雪, 丁倩云, 刘静, 等. 鱼腥草化学成分、药理及质量控制研究进展 [J]. 药物分析杂志, 2022, 42(1): 108-120.
[16]Ji K M, Li M, Chen J J,. Anaphylactic shock and lethal anaphylaxis caused byCordata injection, a herbal treatment in China [J]., 2009, 64(5): 816-817.
[17]刘苗苗, 崔清华, 范路路, 等. 鱼腥草多糖的制备及其体外抗病毒活性研究 [J]. 天然产物研究与开发, 2020, 32(1): 110-117.
[18]黄世琼, 肖礼娥. 药用植物鱼腥草的研究进展 [J]. 现代医药卫生, 2010, 26(19): 2953-2954.
[19]倪俊, 靖恒烨, 高玉婷, 等. 滇西北地区鱼腥草抗氧化活性研究 [J]. 化学与生物工程, 2022, 39(7): 26-31.
[20]Chang Q Y, Lu X, Liu Z,. Identification and characterization of the bZIP transcription factor family in yellowhorn [J]., 2021, 32(1): 273-284.
[21]Yang Y X, Li J W, Li H,. Thegene family in watermelon: Genome-wide identification and expression analysis under cold stress and root-knot nematode infection [J]., 2019, 7: e7878.
[22]Liu J Y, Chen N N, Chen F,. Genome-wide analysis and expression profile of the bZIP transcription factor gene family in grapevine () [J]., 2014, 15(1): 1-18.
[23]Zhao J, Guo R R, Guo C L,. Evolutionary and expression analyses of the apple basic leucine zipper transcription factor family [J]., 2016, 7: 376.
[24]Zhang M, Liu Y H, Shi H,. Evolutionary and expression analyses of soybean basic Leucine zipper transcription factor family [J]., 2018, 19(1): 159.
[25]Jakoby M, Weisshaar B, Dröge-Laser W,. bZIP transcription factors in[J]., 2002, 7(3): 106-111.
[26]Wang W W, Wang Y F, Zhang S M,. Genome-wide analysis of the abiotic stress-related bZIP family in switchgrass [J]., 2020, 47(6): 4439-4454.
[27]徐洪伟, 金妍, 刘美琦, 等. 番茄bZIP转录因子响应低温胁迫的功能研究 [J]. 吉林师范大学学报: 自然科学版, 2023, 44(1): 112-122.
[28]Liu C T, Ou S J, Mao B G,. Early selection of bZIP73 facilitated adaptation ofto cold climates [J]., 2018, 9(1): 3302.
[29]Mirzaei K, Bahramnejad B, Fatemi S. Genome-wide identification and characterization of the bZIP gene family in potato () [J]., 2020, 24: 100257.
[30]Hwang I, Jung H J, Park J I,. Transcriptome analysis of newly classifiedtranscription factors of Brassica rapa in cold stress response [J]., 2014, 104(3): 194-203.
Identification and analysis ofgene of
CAI Wei1, XIAO Jing-zhong3, YU Xiao-li3, YAO Cai-feng3, CHEN Man3, ZENG An-jie3, LI Fu-rong3, CHEN Liao3, LIU Lei3, FU Hong-bo2
1. School of Resources and Environmental Engineering, Mianyang Normol University, Mianyang 621000, China 2. College of Biological and Agricultural Sciences, Honghe University, Mengzi 661199, China 3. College of Life Science & Biotechnology, Mianyang Normol University, Mianyang 621000, China
To lay a foundation for further study ofgene.Based on the transcriptome data of, we identified the HcbZIP family genes and analyzed their physicochemical properties, phylogenetic tree, motif compositions and the expressions in two different ecotype.A total of 163genes were identified. The number of amino acids encoding proteins ranged from 115 aa (and) to 703 aa (). More than 53% of the family members were basic proteins, and more than 84% of the family members were unstable proteins, all members of the family were hydrophilic proteins. In the secondary structure, 161 family members are composed of four parts: alpha helix, extended strand, beta turn and random coil. The ratio of beta turn was the lowest, and the ratio of alpha helix and random coil were the highest. Phylogenetic tree analysis divided these 163 family members into 9 classes, including C, S, G, A, I, U, H, F and D, the largest number was in class D with the number of 35, the lowest number was in classes C with only four, and nogenes were classified into classes E and B. Motif composition analysis revealed that motif 1 (RLLQNRESARRSRLRKKA- YVQELESSVAK) was highly conserved in each family member and constitutes a conserved domain of thegene. The motifs of the genes in different classes were close, and part of the motifs only existed in the genes in specific classes. The expression analysis showed that the number ofgene expressed in 7# was more than that in 6#, it was also found that the number ofgene only expressed in 7# was more than that in 6#. A total of 6genes including,,,,, andwere highly expressed in 6# and 7#. By comparing the differentially expressedgenes of 6# and 7#, we found that a total of 96 differentially expressedgenes, which were divided into 2 classess, and 63 differentially expressed genes were highly expressed in 7#, the expression level of the other 33 differentially expressed genes were higher in 6#.A total of 4 genes, including,,andwere selected as candidate genes that can adapt to different ecological conditions for these two materials.
Houttuynia cordata Thunb.; bioinformatics; differentially expressed gene; different ecotypes; genes family
R286.12
A
0253 - 2670(2023)13 - 4295 - 11
10.7501/j.issn.0253-2670.2023.13.022
2022-12-06
四川省科技计划项目(2021YJ0115);四川省科技计划项目(2020YFN0113)
蔡 伟(1989—),硕士,助教,主要从事环境生物学、植物资源评价与利用研究,Tel: 15983684595 E-mail: 708810337@qq.com
通信作者:刘 雷(1980—),硕士,副研究员,主要从事药用植物资源评价与利用、农业产业化与标准化研究。E-mail: 33020897@qq.com
付鸿博(1987—),博士,讲师,主要从事植物种质资源创新与分子育种。E-mail: 113311168@qq.com
[责任编辑 时圣明]
