多酚类化合物基于非编码RNA调控发挥抗肿瘤及辐射增敏作用研究进展
2023-04-06王小龙郝利民刘永奇刘可可张雪梅鲁吉珂伊娟娟
李 雪,李 俐,王小龙,郝利民,刘永奇,刘可可,张雪梅,鲁吉珂,*,伊娟娟,*
(1.郑州大学生命科学学院,河南 郑州 450001;2.河南省银丰生物工程技术有限公司,河南 郑州 450001;3.军事科学院系统工程研究院,北京 100010)
恶性肿瘤已成为威胁人类健康的重大公共卫生问题,已引起广泛关注[1]。目前放疗是治疗恶性肿瘤常用方法之一,在临床放疗过程中,所采用的辐射粒子通常会引起肿瘤细胞DNA发生不同程度损伤,进而影响DNA完整性或改变其化学结构,最终诱发细胞发生凋亡[2]。放疗对一些辐射敏感性较高的肿瘤有较好的疗效,但多数肿瘤自身特性决定的放疗抗性仍然是干扰临床疗效的一大障碍[3]。因此,在放疗过程中,需要利用辐射增敏剂等辅助手段来提高细胞辐射敏感性[4-5]。目前已陆续开发出多种化学辐射增敏剂,但人工合成辐射增敏剂大多存在不易合成、价格昂贵、毒性较大和靶向性低等缺点,限制了其在辐射增敏方面的应用。从天然活性成分资源中寻找并开发的辐射增敏剂在效用、成本和毒副作用等方面均具有其特殊的优势,是辐射增敏抗肿瘤的有效途径,也是辐射增敏剂开发的重要方向之一。
多酚类化合物是一类植物次级代谢产物,其种类繁多,几乎存在于所有的植物中[6]。多酚类化合物具有良好的电离辐射防护作用,能够有效降低电离辐射造成的各种组织细胞氧化损伤。另外,多酚类化合物具有显著的抗肿瘤活性,因其自身毒性低、无蓄积及疗效好等优势而成为一类极具开发价值的天然抗肿瘤活性成分。已有研究表明,多酚具有多靶点作用,可以介导多种细胞信号通路,对不同的肿瘤细胞均具有辐射增敏作用,进而能提高其放疗效果,因此多酚具有开发为天然辐射增敏剂的潜能[7-8]。目前,非编码RNA(non-coding RNA,ncRNA)已被证实能够调节癌基因或抑癌基因,从而发挥促癌或抑癌作用[9]。值得强调的是,研究发现多酚类物质能够基于ncRNA调控水平发挥其抗肿瘤及辐射增敏抗肿瘤作用[10]。因此,本文对多酚类化合物抗肿瘤及辐射增敏作用及其机制进行综述,并重点突出多酚类化合物基于ncRNA调控层面的作用效果,以期为天然多酚类化合物参与肿瘤等疾病的干预治疗及相关特医膳食食品的开发提供参考。
1 ncRNA与肿瘤及电离辐射的关系
ncRNA是指从DNA转录而来,不具备编码蛋白质能力的RNA,主要包括微小非编码RNA(microRNA,miRNA)、长链非编码RNA(long chain non-coding RNA,lncRNA)和环状RNA(circular RNA,circRNA)等多种已知功能和未知功能的RNA[11]。
1.1 ncRNA与肿瘤
ncRNA作为关键调节因子参与许多肿瘤细胞的发育、分化、增殖、转录、转录后修饰、细胞凋亡和细胞代谢等过程[12]。miRNA可通过与不同信号通路相关miRNA靶点相结合,在转录后调控基因表达[13]。lncRNA的生物学功能主要包括基因甲基化调控、转录激活、细胞周期调控、DNA损伤修复、mRNA翻译调控[14]。lncRNA具有多种发挥调控肿瘤生物学功能的形式(图1):1)lncRNA可以充当miRNA的前体,通过直接生成miRNA而发挥调控作用;2)通过与miRNA竞争结合mRNA非翻译区,充当“竞争性内源RNA(competing endogenous RNA,ceRNA)”;3)能够诱导染色质重塑而调控基因表达;4)与蛋白质相互作用来调控基因表达;5)参与表观遗传调控,可以通过干扰靶分子甲基化及泛素化来调控基因表达[15]。lncRNA调控功能机制繁多,其中ceRNA调控理论已经得到了越来越多的研究认可[16]。如MALAT1基因通过抑制细胞外调节蛋白激酶/丝裂原活化蛋白激酶(mitogen-activated protein kinase,MAPK)和MMP2基因信号传导以及通过抑制miR-155和促进FBXW7的表达进而充当胶质母细胞瘤中的肿瘤抑制因子[17]。Wang Yue等[18]发现lncRNA-ROR可通过竞争内源性RNA作用来连接miRNA网络与一些核心转录因子,使这些转录因子免受miRNA介导的抑制作用,从而在人类胚胎发育过程中发挥重要的功能。lncRNA是胚胎干细胞的一类新调节因子,一些lncRNA可以作为ceRNA,通过与miRNA竞争性结合来调节mRNA,进而降低小鼠胚胎干细胞致死率[19]。
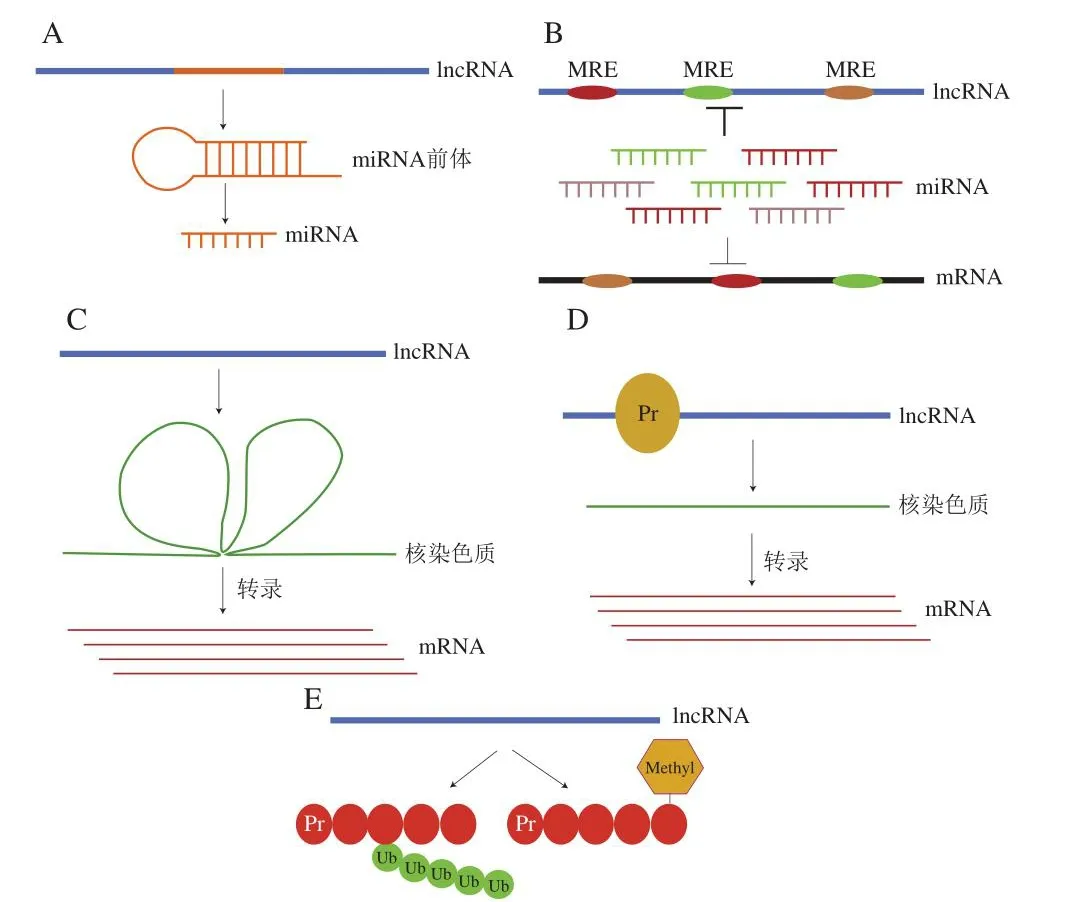
图1 lncRNA调控肿瘤生物学功能的主要作用机制[15]Fig.1 Mechanism by which lncRNA regulates the biological function of tumor cells[15]
此外,circRNA作为环状ncRNA的代表,其分布广泛、细胞功能多样,具有作为新型肿瘤生物标志物和治疗靶标的潜力。circRNA调控肿瘤生物学功能的主要作用形式如下(图2):1)与RNA聚合酶II相互作用并与典型剪接竞争来调节基因转录;2)与RNA结合蛋白(RNA binding proteins,RBP)相互作用;3)在特殊情况下内源性circRNA可编码真核生物中的蛋白质;4)通过作为miRNA的分子海绵影响miRNA介导的靶基因调控和相关细胞信号通路从而发挥抗肿瘤作用[20]。研究表明circ-NFIX海绵化miR-34a-5p通过Notch信号通路上调NOTCH1从而抑制胶质瘤的生长[21]。进一步研究表明circ-SMARCA5可通过海绵化miR-17-3p和miR-181b-5p促进肿瘤抑制因子TIMP3的表达,进而抑制肝癌细胞的增殖和迁移[22]。此外,circRNA在功能上与miRNA相互作用,如circ-0000502/miR-124、circ-0001955/miR-145-5p、circ-0001955/miR-516a-5p和circ-0001955/miR-145-5p均可调控肝癌的发生[23]。
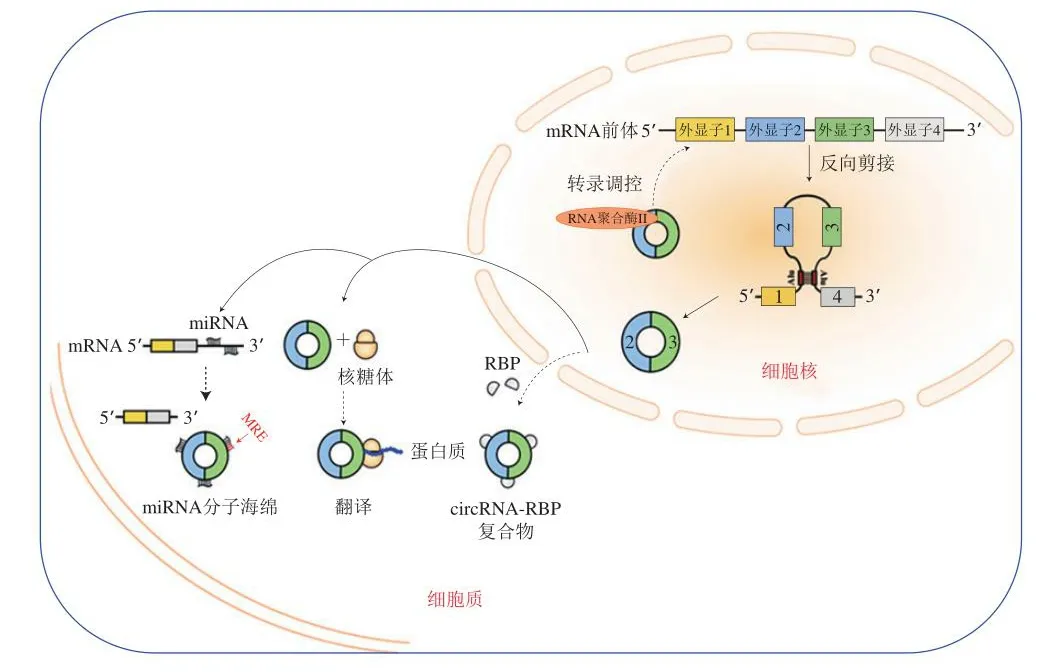
图2 circRNA调控肿瘤生物学功能的主要作用机制[20]Fig.2 Mechanism by which circRNA regulates the biological function of tumor cells[20]
1.2 NcRNA与电离辐射
电离辐射作为治疗癌症的一种有效手段,可以刺激细胞在ncRNA层面上发生胁迫应答。一些miRNA已经被证明能够参与到细胞辐射应答过程中[24-25]。此外,已有研究发现,lncRNA能够通过靶向受辐照细胞中的损伤感应基因、损伤修复基因或凋亡通路相关基因进而调控相应的辐射应答过程[26-27]。已有研究发现,在人外周血单核细胞辐射应答过程中有多种lncRNAs参与,这些差异lncRNAs能够对辐射刺激产生时间依赖式胁迫应答[28]。此外,Aryankalayil等[29]在动物水平上也验证了辐射处理能够调控lncRNA的差异表达,对实验小鼠全身进行不同剂量辐射处理后,提取动物全血总RNA进行深度测序分析,结果显示不同辐射剂量及不同时间处理均会使lncRNA表达模式发生显著变化。利用mRNA/lncRNA微阵列数据结合生物信息学手段在下咽鳞状细胞癌细胞中发现一些辐射抗性相关的lncRNA,这为进一步研究这些lncRNA的作用奠定了基础[30]。UCA1(urothelial cancer associated 1)是1 种在多种肿瘤细胞中异常表达的lncRNA,在具有辐射抗性的前列腺癌细胞中作为功能癌基因异常表达,这与细胞的辐射抗性直接相关。深入的研究发现UCA1可以激活前列腺癌细胞磷脂酰肌醇-3-激酶(phosphoinositide 3-kinase,PI3K)/蛋白激酶B(protein kinase B,PKB/AKT)信号通路,使细胞表现出相应的辐射抗性,进而影响辐射治疗效果[31]。值得注意的是,也有研究表明circRNA同样可以参与到细胞辐射应答过程中,如通过对辐照前后人胚胎肾细胞中的circRNA表达进行高通量测序分析,发现在辐射处理后有158 个circRNAs具有显著差异表达[32]。此外,通过对circRNA-miRNA-mRNA网络调节基因表达的研究发现,电离辐射会影响小鼠骨髓基质细胞中circRNA的表达水平[33]。
2 多酚类化合物基于ncRNA发挥抗肿瘤及辐射增敏作用
植物多酚属于植物次生代谢过程中自然合成的多羟基化合物,主要包括酚酸类、二苯乙烯类、类黄酮类、香豆素类和单宁等[34]。天然产物多酚类化合物因其良好的生物活性而受到科学界的广泛关注[35]。多酚的生物学作用主要在于能够有效抑制活性氧和活性氮的产生,将电子转移给自由基;此外还可以激活抗氧化酶,改善氧化应激和炎症[36-38]。已有研究表明,在多种肿瘤中多酚可基于ncRNA抑制多种肿瘤细胞的增殖、侵袭、迁移等[39-41],其代表性的多酚,如姜黄素、表没食子儿茶素没食子酸酯(epigallocatechingallate,EGCG)、槲皮素、白藜芦醇、咖啡酸苯乙酯、染料木黄酮和葛根素等可通过调节多种信号通路参与抑制肿瘤细胞增殖和诱导其凋亡以及提高相关肿瘤细胞的辐射敏感性[42-43]。
2.1 姜黄素
姜黄素是从姜黄根茎中提取的最具代表性的生物活性多酚,具有多种抗癌特性[44]。研究发现10~40 μmol/L姜黄素可显著降低非小细胞肺癌细胞H460、A549的活力,且呈剂量-效应关系,40 μmol/L姜黄素可将非小细胞肺癌细胞H460、A549的细胞活力降低至30%左右,使细胞凋亡率由5%升高至25%左右;miRNA和circRNA之间存在靶标关联,姜黄素可通过调节circ-PRKCA/miR-384/β1整合素(integrin β1,ITGB1)通路进而抑制恶性肿瘤的发生[45]。此外,姜黄素还可通过上调miR-192-5p/miR-206的表达,抑制PI3K/AKT/mTOR信号通路,进而抑制非小细胞肺癌细胞的增殖和诱导其细胞凋亡[46-47]。研究者通过微阵列对10 μmol/L姜黄素处理的胰腺癌细胞中miRNA表达情况进行分析,结果发现姜黄素可上调miR-22的表达进而抑制其靶基因SP1转录因子和雌激素受体1的表达[48]。在胃癌细胞SGC-7901中,50 μmol/L的姜黄素可显著降低SGC-7901细胞活力并抑制其细胞G0/G1-S期,通过上调SGC-7901细胞中miR-34a的表达,下调Bcl-2、细胞周期蛋白依赖性激酶(cyclin dependent kinase,CDK)4和G1/S-特异性周期蛋白-D1的表达,进而发挥抗癌作用[49]。在人前列腺癌干细胞中,Liu Te等[50]通过细胞实验确定姜黄素对前列腺癌细胞的半数抑制剂量(half-inhibition concentration,IC50)为46.5 μmol/L;同时在动物水平上发现姜黄素能够通过降低lncRNA-ROR表达,相应增加miR-145表达水平,发挥抗肿瘤作用。已有研究表明,姜黄素在乳腺癌细胞中具有调节miRNA表达的潜力,能够影响细胞中致癌miRNAs(miR-19a和miR-19b)和抑癌miRNAs(miR-15a、miR-16、miR-34a、miR-146b-5p和miR-181b)的表达[51]。姜黄素还可通过调控lncRNAMEG3/miR-214及circ-PLEKHM3/miR-320a/SMG1通路进而抑制卵巢癌细胞的增殖并促进其细胞凋亡[52-53]。此外,在肾癌细胞和肝癌细胞中,姜黄素可通过调节circ-FNDC3B/miR-138-5p/胰岛素样生长因子(insulin like growth factor,IGF)2和circ-007871/miR-378b/PRIM2信号通路,发挥抗癌作用[54-55]。
姜黄素对多种恶性肿瘤具有辐射增敏作用,微阵列分析发现miR-1246是姜黄素处理后表达变化最大的miRNA,姜黄素通过调节miR-1246的表达靶向抑制膀胱癌细胞中的p53基因翻译,进而提高膀胱癌细胞对辐射的敏感性[56]。此外,姜黄素可通过激活miR-143抑制其介导的自噬,增强前列腺癌细胞的辐射敏感性[57]。通过建立移植鼻咽癌肿瘤模型鉴定姜黄素的体内辐射增敏作用,结果发现灌胃100 mg/(kgmb·d)姜黄素的小鼠比单独辐照治疗的小鼠体内肿瘤抑制率显著提高,可达到62.18%;通过相关鼻咽癌细胞实验,发现姜黄素可通过上调miR-593的表达,导致多药耐药基因1(multidrug resistance gene 1,MDR1)表达下降,进而促进鼻咽癌细胞的辐射敏感性[58]。同时在鼻咽癌细胞中,通过对姜黄素处理前后circRNA的变化分析,发现姜黄素可通过调节circRNA-miRNA-mRNA,抑制表皮生长因子受体(epidermal growth factor receptor,EGFR)、信号转导和转录激活因子3(signal transducers and activators of transcription 3,STAT3)以及生长因子受体结合蛋白2(growth factor receptor-bound protein 2,GRB2)的表达,从而达到辐射增敏作用[59]。此外,姜黄素还可通过调节circRNA-102115/miR-335-3p/MAPK1的信号通路进而提高鼻咽癌细胞的辐射敏感性[60]。
天然产物姜黄素基于ncRNA调控的相关作用及其机制如表1所示。

表1 天然产物姜黄素基于ncRNA调控的相关作用及其机制Table 1 Related effects and mechanisms of natural curcumin based on non-coding RNA regulation
2.2 表没食子儿茶素没食子酸酯
EGCG是绿茶中的生物活性多酚,可作为一种有效的癌症预防剂和治疗剂[61]。研究发现40 μmol/L的EGCG作用于非小细胞肺癌细胞A549可将G0/G1期细胞比例由55.58%增加到79.15%;通过二代测序分析发现EGCG可影响非小细胞肺癌细胞A549内miRNA表达,从而调节A549细胞周期并抑制其细胞增殖和转移[62]。在宫颈癌细胞中,EGCG能够以剂量和时间依赖性方式显著抑制宫颈癌细胞HeLa生长,在24 h和48 h其IC50分别为90.74、72.74 µg/mL;同时EGCG能够通过调节细胞中相关miRNAs(miR-203、miR-125b、miR-210、miR29)的表达进而抑制不同类型宫颈癌细胞HeLa、SiHa、CaSki、C33A的生长[63]。在乳腺癌细胞MCF-7中,EGCG(5 μg/mL和20 μg/mL)在24~72 h对癌细胞的抑制作用在40%~75%之间,同时5、20 μg/mL的EGCG处理后MCF-7细胞G2/M期从11.00%分别显著增加至20.4%和28.6%,凋亡率分别增长至24%和43%;深入研究揭示EGCG可通过抑制miR-25表达并增加多聚ADP核糖聚合酶(poly-ADP-ribose polymerase,PARP)、含半胱氨酸的天冬氨酸蛋白水解酶(cysteinyl aspartate specific proteinase,Caspase)-3和Caspase-9的表达发挥其抑制乳腺癌细胞增殖的作用[64]。此外,在人肝癌细胞HepG2中,通过微阵列分析发现EGCG处理的人肝癌细胞HepG2中有13 个miRNAs上调,48 个miRNAs下调;研究发现,EGCG可通过上调差异性表达的miR-16,并靶向Bcl-2进而诱导人肝癌细胞HepG2凋亡[65]。此外,EGCG能够通过抑制活性氧和miR-483-3p表达减弱肝癌细胞的转移能力,进而达到抗癌作用[66]。本课题组前期实验表明,茶源多酚类活性组分EGCG、辐射单独处理均可抑制肝癌细胞生长,并且辐射协同EGCG处理抑制效果更加明显,EGCG可进一步激活miR-34a/Sirt1/p53信号通路,进而下调抗凋亡蛋白Bcl-2,上调促凋亡蛋白Bax和Caspase-3的表达,进而提高肝癌细胞辐射敏感性[67]。
天然产物EGCG基于ncRNA调控的相关作用及其机制如表2所示。
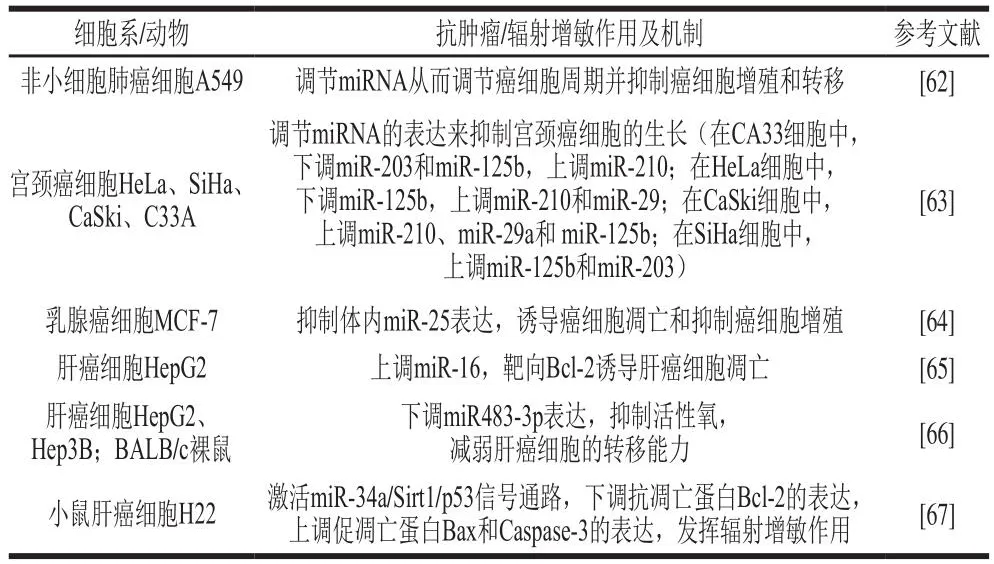
表2 天然产物EGCG基于ncRNA调控的相关作用及其机制Table 2 Related effects and mechanisms of natural EGCG based on non-coding RNA regulation
2.3 槲皮素
槲皮素是一种普遍存在于水果、蔬菜中的类黄酮[68]。槲皮素在对抗包括非小细胞肺癌在内的多种人类肿瘤中发挥重要作用,利用25、50、100 μmol/L的槲皮素作用于非小细胞肺癌GLC-82、HTB-182细胞,结果发现随着槲皮素浓度的增加和处理时间的延长,GLC-82和HTB-182细胞的活力逐渐降低;同时槲皮素可增强Bax和p53的表达,抑制Bcl-2和Chk1的表达,通过上调miR-16-5p的表达,下调WEE1 G2检查点激酶(WEE1 G2 checkpoint kinase,WEE1)的表达增强非小细胞肺癌的辐射敏感性[69]。50 μmol/L槲皮素作用于口腔癌细胞(HSC-6、SCC-9)48 h可将细胞活力降低50%;同时槲皮素可通过调节HSC-6、SCC-9细胞中miR-16和同源盒基因A10(homeobox A10,HOXA10)的表达抑制细胞活力、迁移和侵袭,进而发挥抗癌作用[70]。已有研究表明100 μmol/L槲皮素可将口腔鳞状细胞的活力降低50%左右,并将其凋亡率提升3 倍左右,槲皮素可以通过调节癌细胞中miR-22以及下游通路WNT1/β-catenin的表达从而发挥其抗肿瘤功能[71]。此外,在肺癌细胞、骨肉瘤、卵巢癌细胞和乳腺癌细胞中,槲皮素可通过分别调控miR-16、miR-217、miR-145和miR-146a的表达,进而发挥抗癌作用[72-75]。
天然产物槲皮素基于ncRNA调控的相关作用及其机制如表3所示。
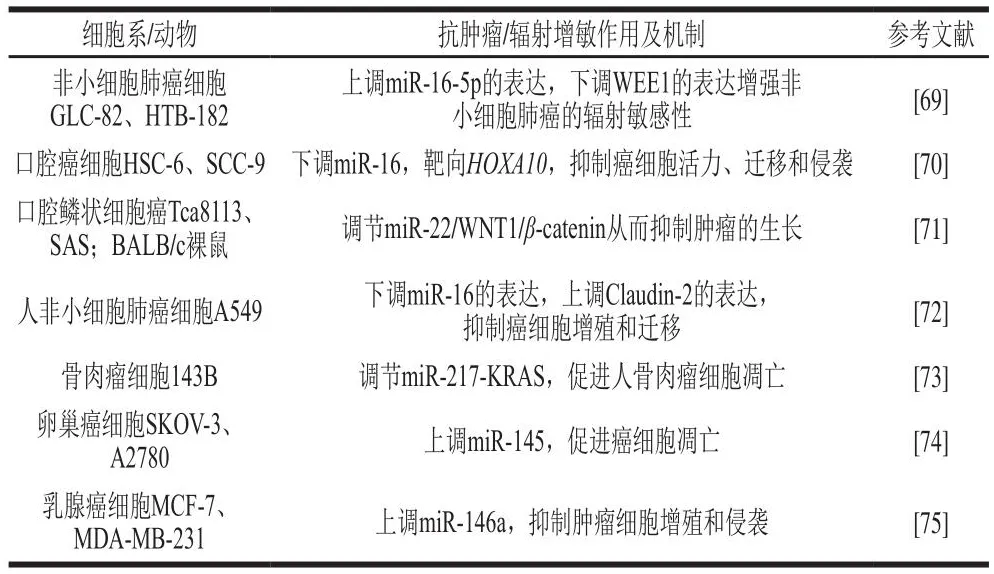
表3 天然产物槲皮素基于ncRNA调控的相关作用及其机制Table 3 Related effects and mechanisms of natural quercetin based on non-coding RNA regulation
2.4 白藜芦醇
白藜芦醇作为天然多酚类化合物已被证明可通过控制细胞生长和分裂、细胞凋亡、转移等多种信号转导途径,影响各种癌症发生及发展的各个阶段[76]。研究发现3.125~100 μmol/L白藜芦醇对非小细胞肺癌细胞(A549和H446)活力的抑制作用呈剂量-效应关系,其IC50为40 μmol/L;通过微阵列分析发现白藜芦醇处理的肺癌A549细胞中有21 个lncRNAs上调,19 个lncRNAs下调,其中AK001796是表达变化最明显的差异lncRNA,是一种促进非小细胞肺癌细胞增殖的致癌基因,白藜芦醇可以下调其表达,进而抑制癌细胞的增殖,发挥抗癌作用[77]。在结肠癌HT-29细胞中,白藜芦醇对HT-29细胞处理48 h的IC50为50.81 μmol/L;值得注意的是,一些相关的lncRNAs(CCAT1、CRNDE、HOTAIR、PCAT1、PVT1、SNHG16)高表达会促进结直肠癌细胞的增殖、迁移和侵袭,白藜芦醇可通过下调这些lncRNAs的表达进而抑制结直肠癌细胞的增殖[78]。此外,白藜芦醇也呈剂量依赖性抑制卵巢癌细胞SKOV-3、OV-90的活力,其对SKOV-3、OV-90细胞的IC50分别为163.31 µmol/L和132.26 µmol/L;此外,白藜芦醇可通过上调细胞中miR-34a的表达直接靶向Blc-2从而发挥抗卵巢癌的作用[79]。也有研究发现藜芦醇还能够通过调节miR-221、miR-17-92、miR-106、miR-17、miR-20、miR-200和miR-122-5p表达来抑制黑色素瘤、前列腺癌、胰腺癌和乳腺癌的发生[80-83]。
天然产物白藜芦醇基于ncRNA调控的相关作用及其机制如表4所示。
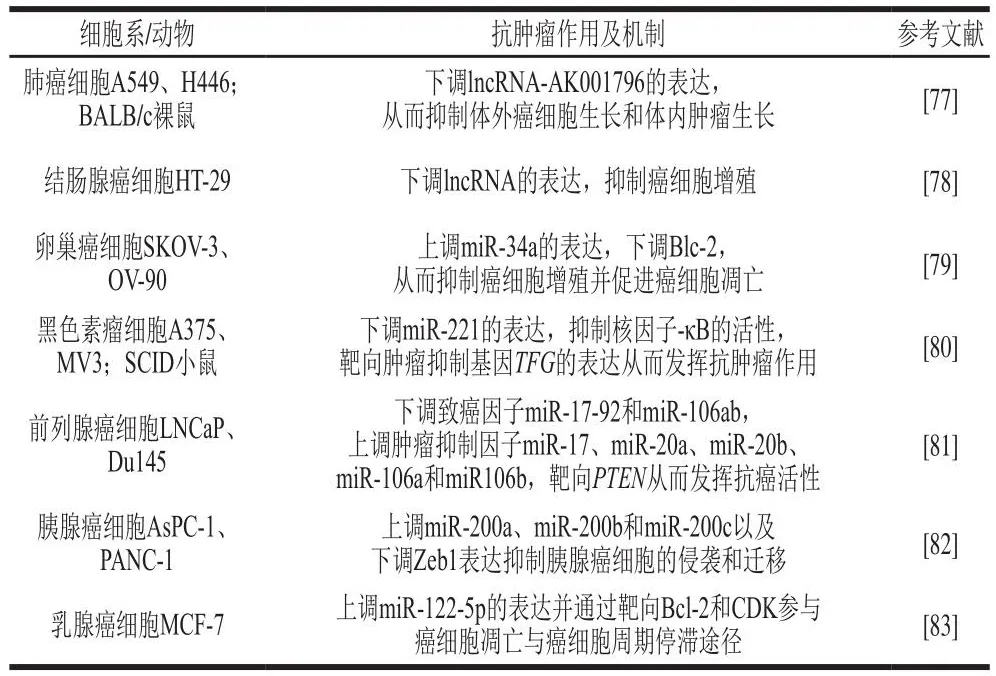
表4 天然产物白藜芦醇基于ncRNA调控的相关作用及其机制Table 4 Related effects and mechanisms of natural resveratrol based on non-coding RNA regulation
2.5 其他多酚类物质
咖啡酸苯乙酯(caffeic acid phenethyl ester,CAPE)是在蜂胶中发现的一种酚类化合物,研究表明CAPE对癌细胞具有很强的抗增殖和细胞毒性,可启动细胞凋亡,减少促炎细胞因子的合成,抑制肿瘤细胞血管新生,增强抗癌药物的作用[84]。本课题组还发现,与单独的CAPE或辐射治疗组相比,CAPE与60Co γ辐射相结合以质量浓度依赖性(1.25~25.00 μg/mL)的方式显著抑制小鼠肝癌细胞增殖,并发挥其辐射增敏潜力,其作用机制与其调控lncRNA表达有关[85]。
染料木黄酮是大豆中的主要异黄酮成分,对不同癌症同样具有抑制作用[86]。在非小细胞肺癌细胞H292、A549中,不同剂量(10~160 μmol/L)的染料木黄酮以时间依赖性抑制H292和A549细胞活力,其IC50在80~160 μmol/L之间;同时染料木黄酮可通过下调circ-0031250的表达,靶向miR-873-5p进而抑制叉头框蛋白(forkhead box,FOX)M1的表达,发挥其调控非小细胞肺癌细胞增殖、迁移、侵袭,促进癌细胞凋亡的作用[87]。在乳腺癌MDA-MB-435和Hs578t细胞中,10~25 μmol/L的染料木黄酮可将其细胞活力降低50%~60%;同时染料木黄酮通过下调miR-155表达,上调FOXO3、PTEN、酪蛋白激酶和p27的表达,在转移性乳腺癌中发挥抗癌作用[88]。此外,染料木黄酮还可通过上调miR-23b的表达抑制乳腺癌的发生[89]。在前列腺癌细胞中,染料木黄酮可通过下调miR-151的表达影响相关靶蛋白的表达抑制癌细胞迁移和侵袭[90]。
葛根素是从葛根的根茎中分离出来的异黄酮衍生物,该活性成分对肺癌、膀胱癌等多种癌症均具有抑制作用[91]。已有研究表明10 μmol/L葛根素可将非小细胞肺癌细胞(A549和H1299)活力降低约50%,并将其细胞凋亡率提高10%以上;通过体内异种移植小鼠肿瘤模型研究发现,葛根素(100 mg/(kgmb·d))治疗能够显著抑制肿瘤生长,同时葛根素可通过上调miR-342抑制体外非小细胞肺癌细胞增殖和体内肿瘤生长,进而发挥抗肿瘤作用[92]。circ-0020394可以通过在葛根素处理的膀胱癌细胞中充当miR-328-3p分子海绵来降低核受体结合蛋白1(nuclear receptor binding protein 1,NRBP1)的表达水平来抑制体内肿瘤发生[93]。以上研究表明天然产物多酚具有调控ncRNA、介导相关通路从而发挥其抗癌功能的潜能(表5)。
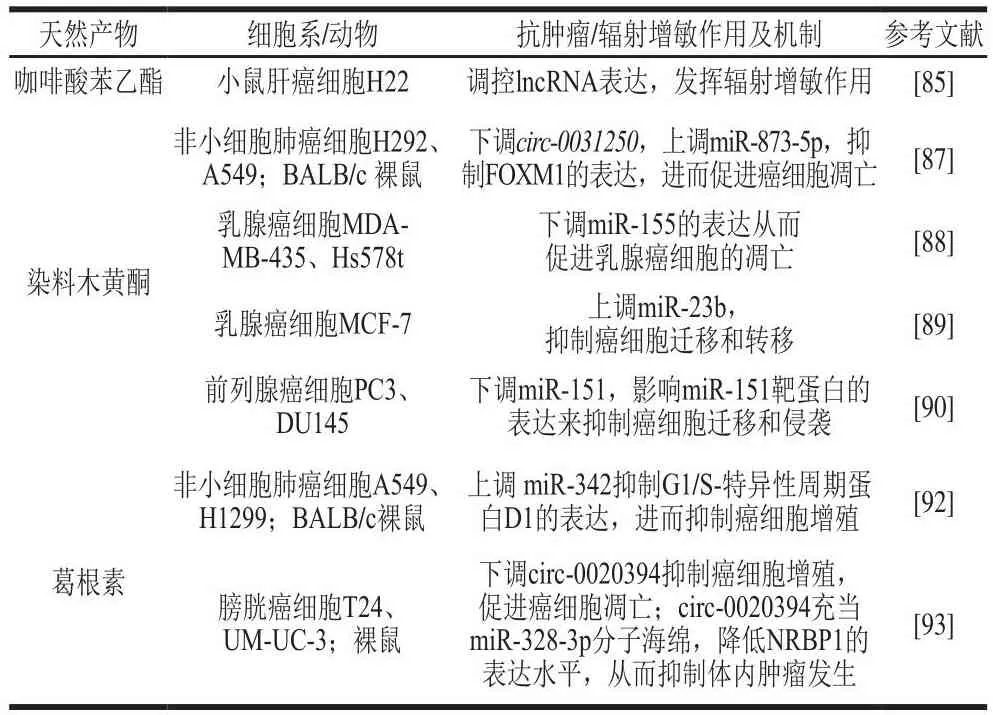
表5 其他多酚基于ncRNA调控的相关作用及其机制Table 5 Related effects and mechanisms of other polyphenols based on non-coding RNA regulation
3 结 语
癌症严重危害人类健康,放疗作为治疗癌症的常用手段存在一定的副作用。多酚类物质可作为辐射增敏剂,在联合放疗治疗癌症方面显示出显著的效果,同时减少电离辐射对正常细胞组织的损伤。ncRNA,尤其是miRNA、lncRNA和circRNA,在调节基因表达方面具有重要的生物学功能。多酚类化合物可通过调控ncRNAs以及影响相关通路的表达进而影响各种癌细胞凋亡、增殖、迁移、侵袭以及增加癌细胞的辐射敏感性。基于此,本文对多酚类化合物抗肿瘤及辐射增敏作用及相应机制进行了综述,并重点探讨了多酚类化合物基于ncRNA调控层面的作用效果,有助于从分子水平明确多酚类化合物抗肿瘤及辐射增敏的治疗靶点,提高其临床应用以及为特医膳食食品开发及应用提供可能性。
然而,尽管已有大量研究表明多酚类化合物是辐射增敏剂的潜在来源,并且多酚类化合物能够靶向癌症相关的ncRNA,特别是调控充当致癌或抑癌的ncRNA从而有效发挥其辐射增敏抗肿瘤作用,为癌症治疗提供了新的干预方法。但仍有如下问题需要关注及解决:1)目前多酚类化合物基于ncRNA调控发挥辐射增敏作用的相关研究仍处于“点状”研究阶段,尚未形成体系,天然产物多酚增敏剂的种类及共性ncRNA靶点或相关信号通路的联系仍是未来研究聚焦的重点。2)鉴于多酚类化合物稳定性差及生物利用度低等问题,未来的研究可以尝试构建绿色纳米递送体系、化学结构修饰及生物合成生产等方式以突破上述方面的限制,提高其应用的范围。3)加强临床试验的推广研究,并在临床试验中证明其有效性及安全性。未来也可以考虑天然辐射增敏剂联合增敏基因治疗的临床应用,并加强多酚联合调控ncRNA系统装载递送体系的开发,增强临床作用效果。
