太平洋牡蛎冷藏过程中闭壳肌品质的变化
2023-04-06杨汝晴张凌晶孙乐常刘光明曹敏杰
欧 韦,杨汝晴,张凌晶,2,孙乐常,2,翁 凌,2,刘光明,2,曹敏杰,2,*
(1.集美大学海洋食品与生物工程学院,福建 厦门 361021;2.大连工业大学海洋食品深加工协同创新中心,辽宁 大连 116034)
牡蛎俗称蚝,是世界上第一大养殖贝类。2020年,我国牡蛎养殖产量达542.5万 t,位居世界首位[1]。牡蛎口感鲜嫩爽滑,富含优质蛋白且营养成分易被机体吸收,深受广大消费者喜爱[2]。但是,牡蛎由于水分含量高、pH值呈中性以及内源性酶活跃等特点,采收后容易发生腐败变质,进而导致营养成分流失和商业价值降低,甚至对人体健康产生潜在威胁[3-4]。
冷藏是最广泛使用的延长牡蛎保质期的手段。但是,随着冷藏时间的延长,牡蛎品质会发生不可逆转的劣化现象。与鱼类相似,贝类在贮藏期间也同样经历软化和腐败,主要是由于其体内化学成分的变化和肌肉蛋白质的降解[5-6]。肌原纤维蛋白是肌肉蛋白的主要组成部分,对于贝类的品质起着至关重要的作用。Yin Zhongzhuan等[7]发现扇贝在冷藏过程中肌原纤维结构出现明显的断裂和卷曲,肌节消失。Xiong Xin等[8]研究发现随着冷藏时间的延长,虾夷扇贝肌球蛋白重链(myosin heavy chain,MHC)逐渐被降解,同时肌肉硬度、弹性和咀嚼性均发生显著下降。水产动物冷藏过程中肌肉蛋白的降解通常是由内源性蛋白酶引起的,如皱纹盘鲍在冷藏过程中丝氨酸蛋白酶和组织蛋白酶L的活力逐渐升高[9]。Liu Bing等[10]发现在4 ℃冷藏过程中丝氨酸蛋白酶抑制剂和半胱氨酸蛋白酶抑制剂能抑制扇贝肌肉中MHC的降解。因此认为丝氨酸蛋白酶和半胱氨酸蛋白酶是导致贝类软化的主要内源性蛋白酶。
目前,国内外对牡蛎的研究主要集中在育种养殖[11]、保鲜技术[12-13]及功能性食品牡蛎肽[14-15]等的方面,关于牡蛎蛋白在冷藏过程中的变化研究较少。随着我国冷链物流业的快速发展以及消费观念的改变,牡蛎冷藏运输量逐年增加,对其在冷藏过程中的品质变化研究也显得更加重要。本研究以太平洋牡蛎为研究对象,分析0 ℃和4 ℃冷藏过程中牡蛎4 个组织(外套膜、腮、内脏团、闭壳肌)中蛋白的降解情况,探究牡蛎在冷藏条件下理化性质指标、质构特性和蛋白降解相关酶(丝氨酸蛋白酶、组织蛋白酶B)活力的变化,重点从闭壳肌蛋白变化的角度揭示牡蛎在冷藏过程中品质变化的机理,旨在为牡蛎冷藏保鲜提供理论参考。
1 材料与方法
1.1 材料与试剂
太平洋牡蛎,2021年10—12月间购于厦门高崎水产市场。去壳后牡蛎肉质量为(19.0±2.0)g。
Boc-Phe-Ser-Arg-MCA、Z-Arg-Arg-MCA荧光底物日本Peptide Institute公司;三羟甲基氨基甲烷(Tris)青岛福林生物化学公司;十二烷基硫酸钠(sodium dodecyl sulphate,SDS)、丙烯酰胺 美国Bio-Rad公司;兔抗人β-肌动蛋白(Actin)抗体 美国Cell Signaling Technology公司;兔抗蓝圆鲹肌球蛋白抗体、兔抗鲍副肌球蛋白(paramyosin,PM)抗体和兔抗扇贝原肌球蛋白(tropomyosin,TM)抗体由本实验制备;辣根过氧化物酶(horseradish peroxidase,HRP)标记的羊抗兔免疫球蛋白G(immunoglobulin G,IgG) 艾比玛特医药科技(上海)有限公司;色谱级无水磷酸二氢钠 上海麦克林公司;色谱级甲醇 赛默飞世尔科技(中国)有限公司;色谱级磷酸 上海阿拉丁生化科技股份有限公司;其他常规试剂如无特殊说明均为分析纯。
1.2 仪器与设备
PT-2100组织捣碎机 瑞士Kinematica公司;Avanti J-26SXP高速冷冻离心机 美国Beckman公司;垂直电泳仪 美国GE Healthcare Bio-Sciences AB公司;G:BOX凝胶成像仪 英国Synegene公司;半干式转移电泳仪大连竟迈生物科技有限公司;成像分析系统 美国Azure Biosystems公司;pH计 上海奥豪斯仪器有限公司;1260液相色谱仪 美国Agilent公司;TA.XT plus/50质构仪 英国SMS公司;FP-8200荧光分光光度计 日本Jasco公司。
1.3 方法
1.3.1 太平洋牡蛎预处理
鲜活太平洋牡蛎去壳,用经过滤的冰海水洗净捞出后沥干。自封袋密封后一半直接置于4 ℃冷库,另一半置于盛有碎冰(0 ℃)的保温箱后,于4 ℃冷库贮存,每2 d更换碎冰。观察表观形态变化。
1.3.2 SDS-聚丙烯酰胺凝胶电泳分析
于第0、1、2、3、4、5、7、9、11天取在0 ℃和4 ℃贮藏的太平洋牡蛎,分离闭壳肌、外套膜、鳃、内脏团4 个组织。加入冰冷(4 ℃贮存)的4 倍体积缓冲液(20 mmol/L Tris-HCl,pH 7.0),组织捣碎2 min。向4 个组织的样品中各加入2 倍体积的蛋白溶解液(20 mmol/L Tris-HCl,pH 7.0,含8 mol/L尿素、1 g/100 mL SDS、体积分数2%β-巯基乙醇),95 ℃加热20 min,用丙烯酰胺质量分数为10%的分离胶和质量分数为5%的浓缩胶对样品进行SDS-聚丙烯酰胺凝胶电泳(polyacrylamide gel electrophoresis,PAGE),分析蛋白条带变化。
1.3.3 闭壳肌Western blotting分析
将第0、1、2、3、4、5、7、9、11天的闭壳肌样品进行SDS-PAGE,蛋白通过半干式电转印转移至硝酸纤维素膜上。将膜以质量分数5%脱脂奶4 ℃过夜封闭,TBST清洗4 次,每次5 min,接着分别用兔抗蓝圆鲹肌球蛋白抗体、兔抗鲍副肌球蛋白抗体、兔抗扇贝原肌球蛋白抗体和兔抗人肌动蛋白抗体孵育1 h,TBST清洗5 次,每次5 min,再用HRP标记的羊抗兔IgG孵育1 h,TBST清洗5 次,每次5 min,加入增强型化学发光(enhanced chemiluminesce,ECL)底物孵育2 min,利用成像分析系统测定。
1.3.4 冷藏过程中相关蛋白降解酶活力的测定
将冷藏不同时间的闭壳肌组织(3 g)捣碎后12 000 r/min离心15 min,上清液即为对应的粗酶液。在900 µL 20 mmol/L Tris-HCl(pH 7.0)缓冲液中加入50 µL粗酶液和50 µL 10 µmol/L的荧光底物,混匀。37 ℃孵育20 min,加入1.5 mL终止液(V(水)∶V(甲醇)∶V(异丙醇)=35∶35∶30)终止反应。采用荧光分光光度计分别在激发波长380 nm、发射波长450 nm条件下测定产物7-氨基-4-甲基香豆素(7-amino-4-methylcoumarin,AMC)的荧光强度。丝氨酸蛋白酶的荧光底物为Boc-Phe-Ser-Arg-MCA、组织蛋白酶B的荧光底物为Z-Arg-Arg-MCA。酶活力单位(U)定义为每分钟释放1 nmol AMC的量。
1.3.5 pH值的测定
参照GB 5009.237—2016《食品安全国家标准 食品pH值的测定》。称取10.00 g绞碎的牡蛎肉,加入100 mL煮沸后冷却的水,组织捣碎,静置30 min后,4 ℃、8 000 r/min离心20 min,取50 mL上清液于100 mL烧杯中,用pH计测定。同一个试样测两次,取平均值。
1.3.6K值的测定
取2.00 g太平洋牡蛎闭壳肌,加入质量分数10%10 mL高氯酸,4 ℃、8 000 r/min离心10 min,向沉淀中加入5 mL质量分数5%高氯酸,均质,再次离心10 min,重复上述操作1 次,合并3 次的上清液,用10 mol/L氢氧化钠溶液调节pH值至6.0,再用1 mol/L氢氧化钠溶液调节pH值至6.4,整个过程在冰上进行。再次离心,超纯水定容至25 mL,过0.22 µm水系膜,置于-80 ℃备用。
高效液相色谱条件[16]:色谱柱SHODEX Asahipak GS-320 HQ;流动相:V(205 mmol/L NaH2PO4)∶V(205 mmol/L H3PO4)=300∶7;流速:0.6 mL/min;柱温:30 ℃;检测波长:260 nm;进样量:20 μL。K值按下式计算。
式中:cATP、cADP、cAMP、cIMP、cHxR、cHx分别为三磷酸腺苷、二磷酸腺苷、腺苷酸、肌苷酸、次黄嘌呤核苷、次黄嘌呤含量/(µmol/g)。
1.3.7 质构特性的测定
将太平洋牡蛎闭壳肌切成高度0.8 cm、直径1.0 cm的肉柱,采用TA.XT plus/50质构仪测定闭壳肌TPA。测定条件:圆柱形探头P/0.5,测定恒定速率1 mm/s,形变50%,两次压缩时间间隔5 s,接触力为5.0 gf。测定的质构参数包括硬度、弹性、咀嚼性和内聚性。每个样品平行测定3 次。
1.4 数据处理与分析
采用SPSS 18.0软件进行单因素分析,利用Duncan检验进行差异显著性分析,所有实验数据均重复3 次进行测定。应用Origin 8.0软件进行绘图。图片采用Adobe Illustrator CS5软件进行标注。
2 结果与分析
2.1 太平洋牡蛎冷藏过程中的表观变化
观察牡蛎的表观变化是检测牡蛎新鲜度最直观的方法。在0 ℃和4 ℃中,太平洋牡蛎的表观变化如图1所示。新鲜太平洋牡蛎闭壳肌呈透明状、组织清晰、肉质紧实。外套膜呈黑色或黑褐色,鳃丝清晰可见,内脏团呈淡黄色或乳白色。在0 ℃贮藏7 d后,牡蛎闭壳肌肉质仍较紧实,冷藏11 d后闭壳肌变成半透明状,肉质变软。在4 ℃贮藏7 d后,闭壳肌色泽变成半透明状,表面渗出较浑浊的黏液,第11天闭壳肌色泽呈不透明状,肉质呈糊状,表面出现黄色黏液。可见,随着冷藏时间的延长,闭壳肌品质变化明显下降,在保鲜效果方面,0 ℃贮藏明显优于4 ℃贮藏。
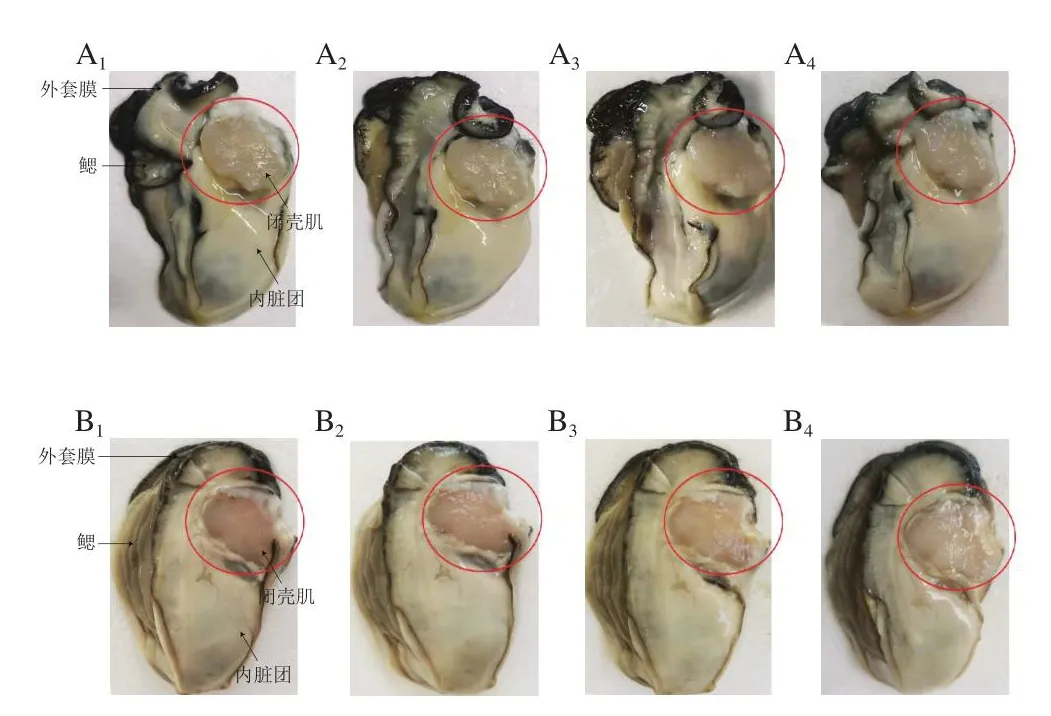
图1 太平洋牡蛎冷藏过程中的表观变化Fig.1 Appearance changes of Crassostrea gigas during cold storage
2.2 太平洋牡蛎冷藏过程中全蛋白SDS-PAGE分析结果
通过SDS-PAGE得到太平洋牡蛎在0 ℃和4 ℃冷藏过程中全蛋白的变化情况。盐溶性蛋白是牡蛎等贝类蛋白质的重要组成部分,主要由肌原纤维蛋白组成[17]。肌原纤维蛋白主要包括肌球蛋白(myosin)、PM、TM和肌动蛋白(actin)。由图2可知,太平洋牡蛎闭壳肌主要含有MHC和Actin,在0 ℃冷藏期间牡蛎组织全蛋白无明显变化。由图3A可知,在4 ℃冷藏期间,闭壳肌在100~150 kDa之间的蛋白条带逐渐出现降解,至第4天,37 kDa处左右出现降解条带。而外套膜、鳃和内脏团无明显降解条带(图3B~D)。表明太平洋牡蛎在4 ℃冷藏过程中,主要是闭壳肌中的蛋白发生了降解。
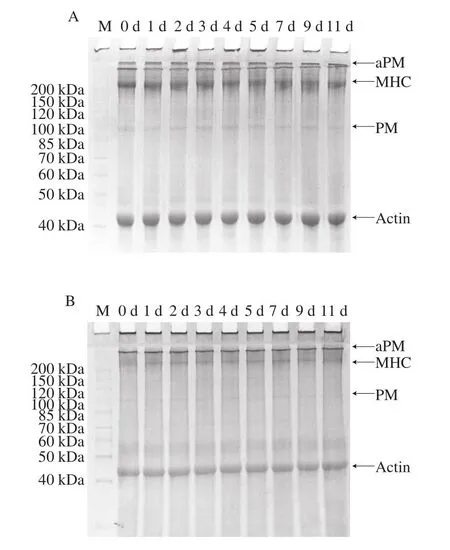

图2 太平洋牡蛎0 ℃冷藏期间蛋白变化Fig.2 Protein changes in Crassostrea gigas stored at 0 ℃
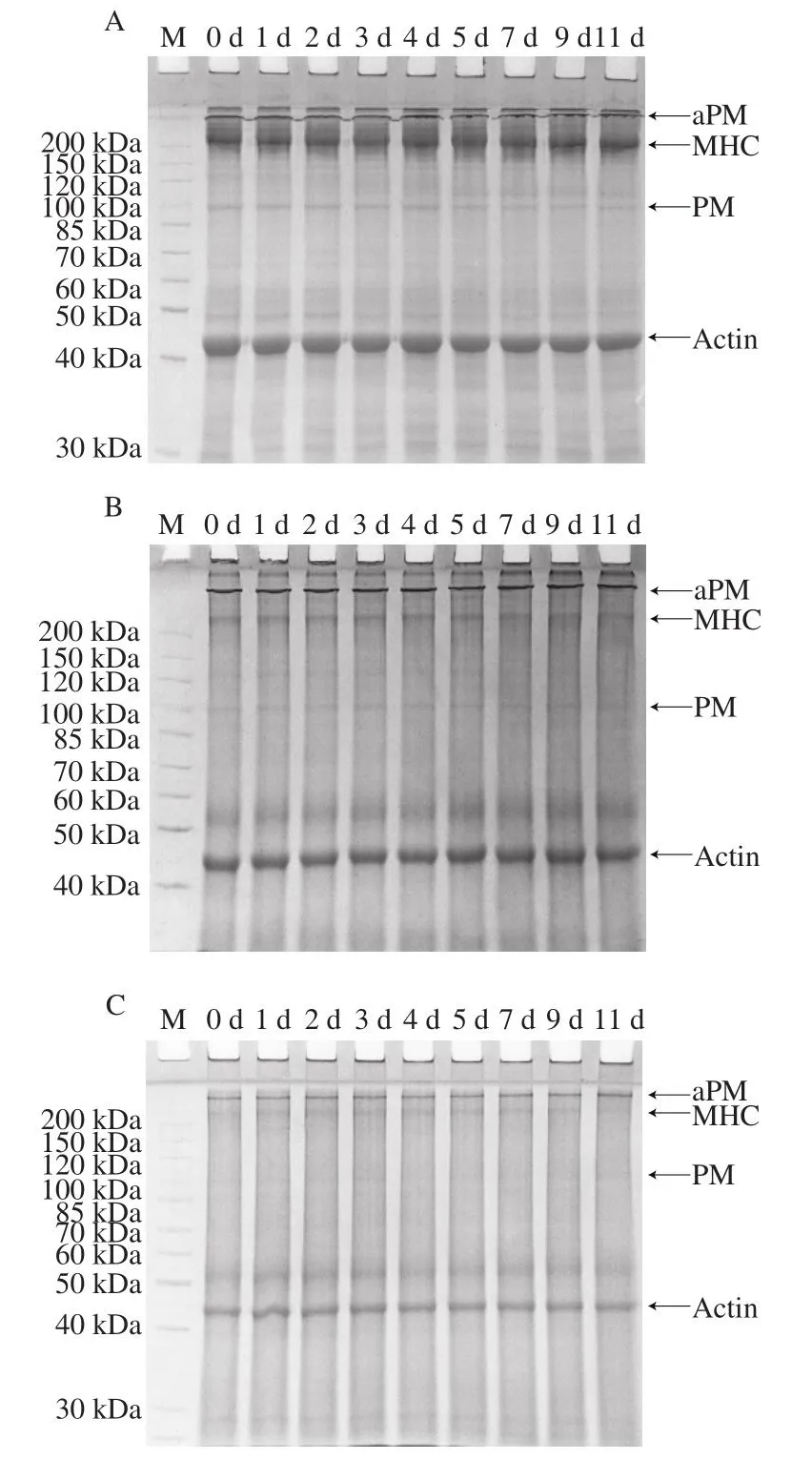

图3 太平洋牡蛎4 ℃冷藏期间蛋白变化Fig.3 Protein changes in Crassostrea gigas stored at 4 ℃
2.3 太平洋牡蛎闭壳肌Western blotting分析结果
为了进一步验证在0 ℃和4 ℃冷藏期间闭壳肌中蛋白的降解情况,采用抗MHC、PM、TM和Actin的多克隆抗体进行Western blotting验证。如图4所示,0 ℃冷藏期间,闭壳肌MHC显色条带为两条,分别在220 kDa和130 kDa左右,后者可能是牡蛎采收后MHC的降解产物。PM主要以多聚体的形式(aPM)存在,单体PM分子质量在97 kDa左右。太平洋牡蛎肌肉蛋白中有两个蛋白与TM抗体发生了免疫交叉反应,分子质量为40 kDa左右,Actin的分子质量为45 kDa左右。太平洋牡蛎在0 ℃贮藏时,闭壳肌MHC在第3天120 kDa处出现降解条带(图4A),而PM、TM和Actin无明显变化(图4B~D)。由图5可知,在4 ℃冷藏过程中,闭壳肌MHC在第1天120 kDa处出现轻微降解条带,冷藏5 d后130 kDa左右处的条带被完全降解,并在110 kDa处出现新的降解条带(图5A),而PM、TM和Actin无明显变化(图5B~D)。表明在4 ℃冷藏时,闭壳肌中MHC最先发生降解,对应SDS-PAGE结果。在冷藏过程中,太平洋牡蛎肌肉蛋白主要是MHC发生降解。
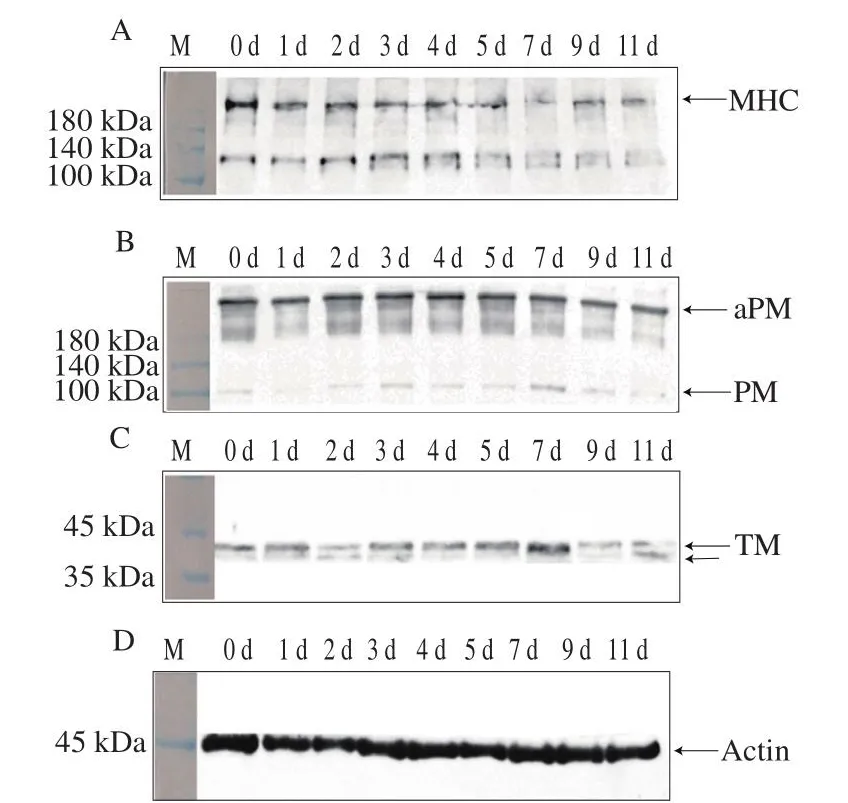
图4 0 ℃冷藏期间太平洋牡蛎闭壳肌蛋白Western blotting分析Fig.4 Western blotting analysis of adductor muscle protein in Crassostrea gigas stored at 0 ℃

图5 4 ℃冷藏期间太平洋牡蛎闭壳肌蛋白Western blotting分析Fig.5 Western blotting analysis of adductor muscle protein in Crassostrea gigas stored at 4 ℃
2.4 太平洋牡蛎闭壳肌冷藏过程中蛋白降解相关酶活力的变化
研究发现,蛋白酶对结构蛋白的降解导致肌肉质地劣化,丝氨酸蛋白酶主要降解肌原纤维和结缔组织蛋白[10],组织蛋白酶B也参与肌肉蛋白质的降解[18]。采用荧光光谱对冷藏期间太平洋牡蛎丝氨酸蛋白酶和组织蛋白酶活力进行测定,结果如图6所示。在新鲜状态下,丝氨酸蛋白酶和组织蛋白酶B的活力均能被检测到,初始酶活力分别为0.11 U/g和0.04 U/g。在0 ℃和4 ℃冷藏前、中期,丝氨酸蛋白酶活力总体呈现先升高后降低的趋势,分别至第3天和第2天达到最大值。贮藏后期(第9天),丝氨酸蛋白酶活力升高可能是微生物的作用,但此时牡蛎已经超出可食用范围。在0 ℃和4 ℃贮藏过程中,组织蛋白酶B的活力也分别在第3天和第2天达到最高。
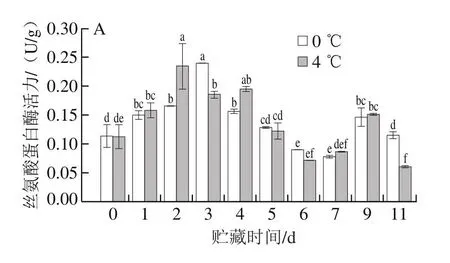
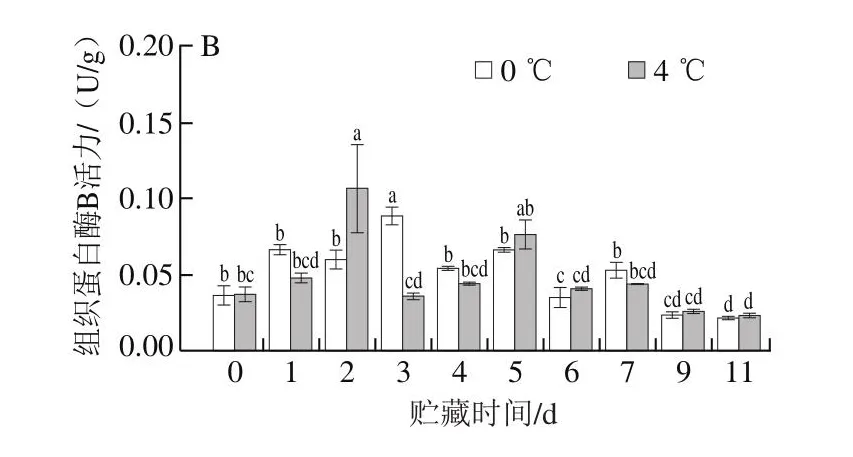
图6 太平洋牡蛎冷藏期间闭壳肌丝氨酸蛋白酶(A)和组织蛋白酶B(B)活力的变化Fig.6 Changes in serine protease (A) and cathepsin B (B) activities in the adductor muscle of Crassostrea gigas during cold storage
2.5 太平洋牡蛎冷藏过程中的理化性质
如图7A所示,新鲜太平洋牡蛎pH值为6.79。在0 ℃冷藏11 d后,pH值为6.52。在4 ℃贮藏5 d后pH值明显下降,到11 d下降至5.84。K值常作为评价食品鲜度的指标,在20%以下为新鲜,20%~40%为二级鲜度,可用于食品加工[19]。新鲜太平洋牡蛎K值为2.51%,冷藏过程K值不断增大。如图7B所示,在0 ℃贮藏时,牡蛎品质劣变较缓慢,在第7天时K值达到22.61%,在第9天时升至40.90%;在4 ℃贮藏3 d后太平洋牡蛎K值达到20.10%,至第5天时,K值为41.83%。从K值的变化可得出:太平洋牡蛎在0 ℃下贮藏的保鲜期比在4 ℃下贮藏更长。
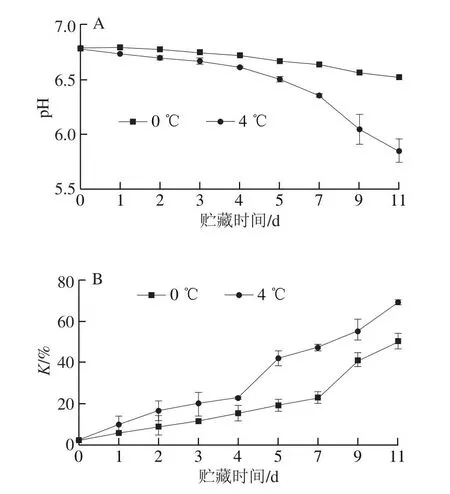
图7 太平洋牡蛎冷藏过程中pH值(A)和K值(B)的变化Fig.7 Changes in pH (A) and K value (B) of Crassostrea gigas during cold storage
2.6 太平洋牡蛎闭壳肌冷藏过程中质构特性的变化
质构是对产品结构或内部构造的感官解释和表达,与产品对压力和触觉属性的反应有关。因此,质构特性常用来表示一些机械特性,如硬度、弹性、咀嚼性和内聚性等[20]。包括肌原纤维蛋白和结缔组织在内的结构蛋白与这些肌肉质构特性相关,因此肌肉的质构特性可用于评估产品的品质[10]。根据表1和表2可知,在0 ℃和4 ℃冷藏过程中硬度和咀嚼性呈先上升后下降的趋势,均在第1天达到峰值,随后逐渐下降,弹性不断下降。由表1可得,在0 ℃冷藏期间,硬度和咀嚼性在第3天恢复到接近新鲜值,冷藏11 d后硬度、弹性、咀嚼性和内聚性分别下降25.5%、2.5%、35.3%和13.2%。根据表2可得,在4 ℃冷藏期间,硬度、弹性和咀嚼性的下降程度均比在0 ℃冷藏期间大,冷藏11 d后硬度、弹性、咀嚼性和内聚性较新鲜组分别下降47.4%、13.7%、51.5%和13.2%。以上结果表明,太平洋牡蛎闭壳肌质地在冷藏期间发生了明显的劣化,表现为硬度、弹性和咀嚼性的下降。
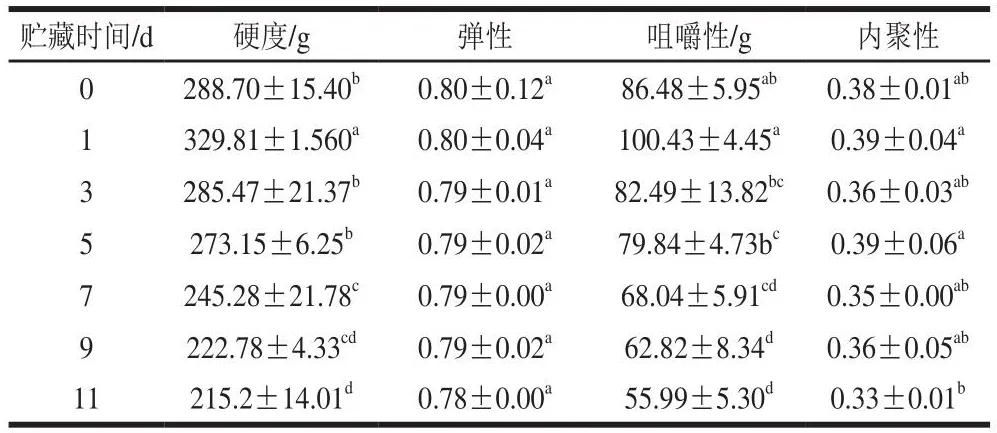
表1 0 ℃冷藏下太平洋牡蛎闭壳肌的质构特性变化Table 1 Textural changes in Crassostrea gigas adductor muscle stored at 0 ℃
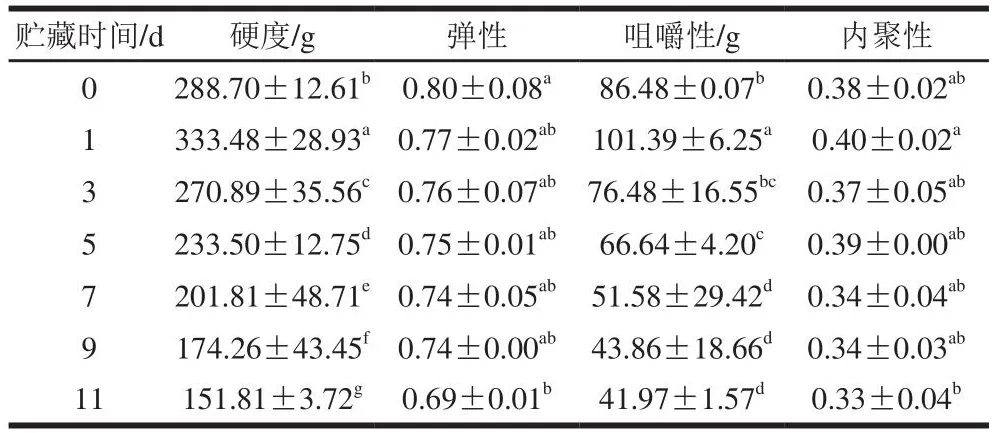
表2 4 ℃冷藏下太平洋牡蛎闭壳肌的质构特性变化Table 2 Textural changes in Crassostrea gigas adductor muscle stored at 4 ℃
3 讨 论
作为牡蛎生产大国,我国用于加工的牡蛎量仅占总产量的30%~40%,大部分仅采用冷鲜初级加工[21]。与鱼类一样,牡蛎在冷藏过程中不可避免地会发生鲜度降低和品质劣化,其主要原因是体内发生一系列生化变化,导致肌肉蛋白质被分解成一系列的中间产物和可溶性含氮物,最终导致微生物大量繁殖,失去食用价值[22]。牡蛎可食用软体部分主要是由闭壳肌、外套膜、鳃和内脏团组成,而闭壳肌总蛋白质含量显著高于外套膜、鳃和内脏团[23]。本研究将太平洋牡蛎肉在0 ℃和4 ℃下进行冷藏,结果显示随着贮藏时间的延长,闭壳肌变化最明显,肉质逐渐变软,边缘和表面开始变得模糊。同时通过对闭壳肌、外套膜、鳃和内脏团全蛋白进行分析,结果显示4 个组织中闭壳肌中全蛋白有轻微分解。因此,牡蛎冷藏过程中主要是闭壳肌蛋白发生了降解,通过对闭壳肌的品质变化分析可作为评价牡蛎肉鲜度的指标。
牡蛎闭壳肌肌肉组成中以肌原纤维蛋白为主,肌原纤维蛋白是一种重要的结构蛋白,由细肌丝和粗肌丝构成,正常生理条件下两种肌丝通过相互滑动来完成肌肉的伸缩运动[24]。细肌丝主要由Actin和TM组成。在无脊椎动物中,粗肌丝由PM和Myosin组成[25]。通过Western blotting检测在0 ℃和4 ℃冷藏过程中MHC、PM、TM和Actin的变化,发现太平洋牡蛎闭壳肌中的MHC分别在第3天和第1天发生降解。海湾扇贝在冷藏过程中也有同样的规律,随着冷藏时间延长MHC逐渐发生降解[26]。牡蛎TM的含量较低,通过SDS-PAGE蛋白条带无法观察到。通过Western blotting检测TM,发现其呈两条带,分子质量大约为40 kDa,与张凌晶等[27]关于太平洋牡蛎粗提TM的研究结果一致。水产品肌肉的K值和pH值与其新鲜程度密切相关。K值常作为评价水产品鲜度的指标。本研究发现,太平洋牡蛎在0 ℃和4 ℃贮藏过程中K值分别在第9天和第5天超过二级鲜度范围。同时,冷藏过程pH值略微降低。由于机体的糖原被降解生成乳酸等酸性物质,造成肌肉的pH值下降,下降程度和肌肉中糖原含量有关,而贝类中糖原含量丰富[28]。
内源性蛋白酶对肌原纤维蛋白和细胞骨架蛋白的水解是水产品贮藏过程中肌肉软化的主要原因[29]。闭壳肌硬度、咀嚼性在第1天达到峰值,随着贮藏时间的延长结构蛋白被内源性蛋白酶降解,纤维间的间隙增大,结构疏松,导致硬度、弹性、咀嚼性和内聚性下降[30-31]。在贝类中,丝氨酸蛋白酶和组织蛋白酶与肌肉降解有关。游银川等[9]发现皱纹盘鲍在冷藏过程中丝氨酸蛋白酶和组织蛋白酶活力逐渐升高,且纯化的组织蛋白酶对肌球蛋白重链有一定的降解作用。任秋颖等[32]从虾夷扇贝中纯化的丝氨酸蛋白酶能显著降解肌球蛋白重链。在0 ℃和4 ℃冷藏前期,太平洋牡蛎闭壳肌中丝氨酸蛋白酶和组织蛋白酶B的活力呈现先升高后下降的趋势,这与皱纹盘鲍[9]和虾夷扇贝[32]中酶活力变化情况一致。组织蛋白酶B在闭壳肌中的活力较低,本实验室前期研究发现太平洋牡蛎内脏团中组织蛋白酶B和L的活力较高,且组织蛋白酶L只在内脏团中表达[33]。冷藏前期蛋白酶(丝氨酸蛋白酶和组织蛋白酶B)活力逐渐上升,可能是酶原在贮藏期间被激活,进而降解肌肉蛋白。冷藏后期蛋白酶活力上升可能是受到微生物的影响。综上所述,牡蛎在0 ℃和4 ℃冷藏过程中内源性蛋白酶在肌肉软化中发挥重要作用,可使肌原纤维蛋白发生降解,从而导致肌肉软化。
4 结 论
在冷藏过程中,太平洋牡蛎闭壳肌中肌肉蛋白发生了降解,且主要降解蛋白为MHC。丝氨酸蛋白酶和组织蛋白酶的活性在贮藏初期逐渐升高,表明这些酶可能参与闭壳肌肌肉组织中MHC的降解,导致闭壳肌的软化以及蛋白质成分降解,可能导致营养价值下降。同时,随着贮藏时间的延长,闭壳肌中pH值逐渐降低、K值逐渐增加,且质构特性也随之劣变。
