侵染广西辣椒的两种单组分菜豆金色花叶病毒属病毒的分子特征
2023-03-13李金哲汤亚飞莫翠萍罗婉笛陈锦清蔡健和邓清超章松柏李战彪
李金哲,汤亚飞,莫翠萍,罗婉笛,陈锦清,蔡健和,邓清超,章松柏,李战彪
(1.农林病虫害预警与调控湖北省工程技术研究中心/长江大学,湖北 荆州 434025;2.广西壮族自治区农业科学院植物保护研究所/农业农村部华南果蔬绿色防控重点实验室/广西作物病虫害生物学重点实验室,南宁 530007;3.广东省农业科学院植物保护研究所/广东省植物保护新技术重点实验室,广州 510640)
【研究意义】辣椒(CapsicumannuumL.)是重要的蔬菜作物,年播种面积在1.5×106~2.0×106hm2,辣椒产业已成为蔬菜的第一大产业[1-2]。随着辣椒种植面积和种植区域的逐步扩大,辣椒病毒病日益严重。因此,明确侵染辣椒的病毒种类及病毒分离物间的遗传进化关系有助于辣椒病毒病的防控,同时为辣椒抗病毒育种提供理论指导。【前人研究进展】世界上已报道侵染辣椒的病毒有70种以上[3-5],国内报道33种[6-9]。由于气候、环境等因素影响,不同地区辣椒感染的病毒种类也有差异,王少立等[10]发现侵染山东辣椒的主要病毒为黄瓜花叶病毒(Cucumber mosaic virus,CMV)和辣椒轻斑驳病毒(Pepper mild mottle virus,PMMoV),汤亚飞等[11]通过对广东省辣椒病毒种类进行检测发现,侵染广东辣椒的优势病毒为辣椒轻斑驳病毒(PMMoV)、甜椒脉斑驳病毒(Pepper veinal mottle virus,PVMV)和辣椒脉斑驳病毒(Chilli veinal mottle virus,ChiVMV)。付尚松[12]对贵州辣椒病毒病调查检测中发现,黄瓜花叶病毒(CMV)、烟草花叶病毒(Tobacco mosaic virus,TMV)和辣椒轻斑驳病毒(PMMoV)为侵染辣椒的主要病毒。严婉荣等[13]在对侵染海南省辣椒的病毒种类进行调查检测时发现,侵染海南辣椒的优势病毒为黄瓜花叶病毒(CMV)和烟草花叶病毒(TMV)。龚明霞等[14]则证实侵染广西辣椒的优势病毒为小米椒内源 RNA 病毒 1(Capsicumfrutescens endornavirus 1,CFEV 1)、辣椒脉斑驳病毒(ChiVMV)、黄瓜花叶病毒(CMV)和甜椒脉斑驳病毒(PVMV)。近年来,双生病毒侵染辣椒的现象也日益严重,范晓堃[15]在西南地区采集辣椒样品进行检测,发现样品中存在番茄黄化曲叶病毒(Tomato yellow leaf curl virus,TYLCV)。刘晨等[16]在对陕西不同地区辣椒病毒进行调查检测时发现,各地区辣椒均携带TYLCV,胡明鑫等[17]在天津辣椒样品中鉴定出TYLCV。迄今为止,尚未有双生病毒侵染广西辣椒的相关报道。【本研究切入点】近年来,广西农科院植保所植物病毒课题组持续关注茄科作物病毒病的发生情况,2022年3月,该课题组在调查中发现,广西百色市的辣椒叶片呈现皱缩、黄化、叶片上卷等症状,疑似受到双生病毒侵染,但具体病原种类尚不明确。【拟解决的关键问题】拟通过分子生物学手段,利用PCR、分段克隆、序列比对和进化树构建等方法对疑似感染双生病毒的辣椒样品进行鉴定,并对各病毒分离物进行遗传进化分析,研究结果将为辣椒病毒病的高效防控提供理论依据。
1 材料与方法
1.1 样品来源
毒源:2022年采自广西百色田间出现黄化、皱缩、上卷等症状的辣椒叶片样品(图1),编号为BS64、BS65、BS66、BS67,样品于-80 ℃保存备用。
1.2 辣椒总DNA提取
取100 mg辣椒叶片样品,选择上海生工生物工程股份有限公司的Ezup 柱式植物基因组 DNA 提取试剂盒进行总DNA提取,具体操作按试剂说明书进行,最终加入50 μL去离子水溶解DNA沉淀,将得到的DNA溶液置于-20 ℃冰箱中保存备用。
1.3 广西辣椒样品中菜豆金色花叶病毒属病毒的PCR检测
利用菜豆金色花叶病毒属病毒通用简并引物AV494:GCCYATRTAYAGRAAGC,CMAG/CoPR:GA NGSATGHTRCADGCCATATA[18-19]对采集到的辣椒叶片总DNA进行PCR检测,以辣椒叶片总DNA为模板,对4份疑似病样进行PCR检测。反应体系(25 μL):2×DreamTaqPCR Master Mix[生工生物工程(上海)股份有限公司]12.5 μL,10 μmol/L AV494和CoPR 引物各 0.5 μL,辣椒叶片总DNA 1 μL,灭菌水10.5 μL。反应程序:95 ℃ 3 min;95 ℃ 30 s;52 ℃ 45 s;72 ℃ 30 s,35 个循环;72 ℃ 10 min。所得的PCR 产物经1%琼脂糖凝胶进行电泳检测,检测为阳性的辣椒样品保存备用。
1.4 广西辣椒样品中菜豆金色花叶病毒属病毒全基因组序列扩增
为获得侵染辣椒的双生病毒的全基因序列,参考已登录GenBank的TYLCV(GenBank登录号:MG904859和KY783940)、中国番木瓜曲叶病毒(Papaya leaf curl China virus, PaLCuCNV)(GenBank登录号:KU892661和MW779523)设计2对背靠背引物,TYLCV-F:TTGGTGGGCCCTCTGGAATG/TYLCV-R:TAACTGTAGCATGAAATTTCCTCATC,PaLCuCNV-F:ACTGTTACTGGTGGTCAGTATGCTAGT/Pa LCuCNV-R:GCGTTTCTTAAGAGTATTTAGGG。以辣椒叶片总DNA为模板进行PCR扩增,PCR 反应程序:95 ℃ 3 min;95 ℃ 15 s,52 ℃ 45 s,72 ℃ 3 min,35 个循环;72 ℃ 10 min。反应体系(50 μL):DNA模板2 μL,正反向引物(10 μmol/L)各0.5 μL,2×Phata Max Master Mix 25 μL,ddH2O 22 μL。
将PCR扩增获得的靶标条带利用DNA凝胶回收试剂盒[生工生物工程(上海)股份有限公司]进行分离纯化。纯化后的PCR产物克隆至pMD18T载体得到连接产物,将得到的连接产物转化大肠杆菌感受态细胞DH5α,获得的菌液涂布含有Amp抗生素的LB培养基平板,平板倒置培养过夜。委托生工生物工程(上海)股份有限公司对确定为阳性克隆的菌落进行测序。
1.5 广西辣椒样品中菜豆金色花叶病毒属病毒的序列分析及进化树构建
利用SeqMan软件对从辣椒样品中扩增获得的序列进行拼接,获得病毒基因组全长序列,将获得的病毒全基因序列在NCBI中利用在线软件ORF finder 和blastn分别进行ORF预测和序列比对分析。进一步利用SDT软件对获得的病毒基因组全长序列进行核苷酸相似性比对分析;通过MEGA 7.0软件进行进化树的构建,选择最大似然法为最优树,步值设置为1000。
2 结果与分析
2.1 辣椒叶片样品的 PCR 检测结果
采用菜豆金色花叶病毒属病毒的简并引物 AV494/CoPR对辣椒叶片样品的总 DNA进行PCR检测, 健康的辣椒叶片样品为阴性对照,确定感染TYLCV的番茄叶片样品为阳性对照。PCR检测结果显示,4份样品均可扩增出长度为570 bp的特异目的片段,而健康的辣椒叶片未扩增出任何条带。

图1 田间被双生病毒侵染的辣椒叶片症状Fig.1 Symptoms of the leaves of pepper infected by begomoviruses in filed
将所得的PCR产物直接进行测序,所得序列在GenBank中进行blastn分析,结果发现,所得序列分别与TYLCV、PaLCuCNV等病毒分离物具有较高的核苷酸相似性。以上结果证实4份辣椒样品中均存在菜豆金色花叶病毒属病毒。
2.2 广西辣椒样品中菜豆金色花叶病毒属病毒分离物基因组克隆及序列分析
为获得辣椒中菜豆金色花叶病毒属病毒全长基因组,从检测为阳性的辣椒总DNA中挑选2个样品(编号:BS66、BS67)分别进行PCR扩增,以辣椒叶片总DNA为模板,参考已登录GenBank的TYLCV(GenBank登录号:MG904859和KY783940)、PaLCuCNV(GenBank登录号:KU892661和MW779523)设计2对背靠背引物:TYLCV-F/TYLCV-R、PaLCuCNV-F/PaLCuCNV-R。对检测为阳性的2个辣椒样品分别扩增后经琼脂糖凝胶电泳分析(图2)发现,BS66利用2对引物均可扩增出约2.7 kb的PCR条带,而BS67仅TYLCV-F/R可以扩增出约2.7 kb的PCR条带。切取PCR扩增获得的靶标条带进行分离纯化、连接、转化等,经菌落PCR检测后,挑选阳性克隆进行测序。经克隆、序列测定和BLAST比对分析发现,从辣椒样品BS66、BS67中共获得3条全基因序列,分别为BS66-1、BS67-1、BS66-2。其中,BS66-1与TYLCV分离物的核苷酸相似性均在91%以上,BS67-1与大部分用于分析TYLCV分离物的核苷酸相似性均在91%以上,但与TYLCV韩国、伊拉克、西班牙分离物(GenBank登录号:ON982201、MT583814、AF071228)则在90%以上。BS66-1与番茄黄化曲叶病毒中国分离物(GenBank登录号:MT969010)相似性最高在99.67%,与TYLCV中国北京、广东、福建等地分离物(GenBank登录号:KT338296、MW779534、MN842311)的相似性为92.69%、92.86%、92.79%,与TYLCV墨西哥分离物(GenBank登录号:EF523478)相似性也在92%以上。BS67-1与TYLCV广西分离物(GenBank登录号:MW389934)相似性最高达98%,与TYLCV中国山西、广东、辽宁等地的分离物(GenBank登录号:OM677619、MW735449、KJ754191)相似性达93.98%、91.84%、91.88%。序列BS66-2与PaLCuCNV广西分离物(GenBank登录号:MW779523、JX12 8102)相似性最高达99.67%,其中与中国重庆、河南分离物(GenBank登录号:OL310479、EU8 74386)相似性达92.49%、95.68%,与越南分离物(GenBank登录号:KC878474)相似性达92.06%。
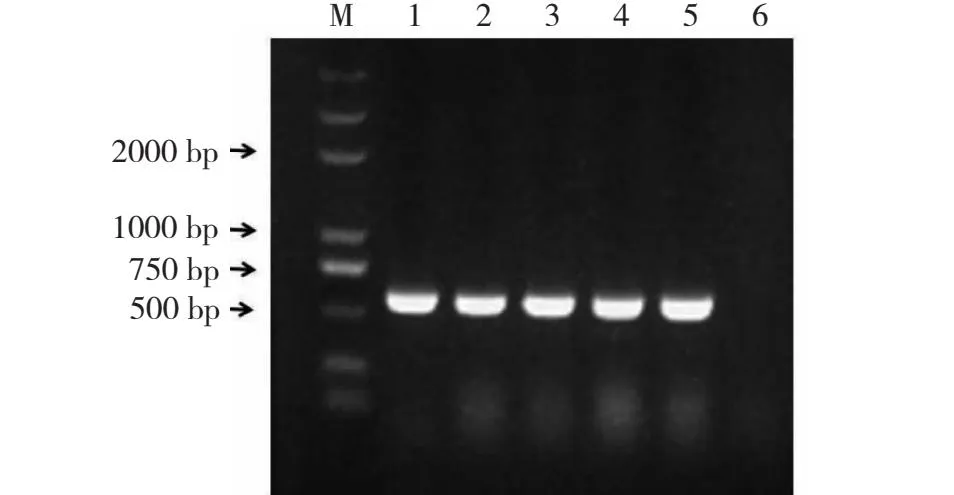
M.DL 2000 DNA标记;1.阳性对照;2~5.样品BS64-67;6.CK(健康对照样品)。M.DL 2000 DNA Marker(TaKaRa);1.Tomato leaf sample infected by TYCD(CK+);2-5.Diseased pepper leaf samples;6.CK(Control sample).
使用SDT软件对获得的3条全基因序列进行核苷酸相似性分析,发现TYLCV广西辣椒分离物BS66-1、BS67-1与本研究用于分析的其他TYLCV各分离物的核苷酸相似性大部分在91%以上(图3)。PaLCuCNV广西辣椒分离物BS66-2与本研究用于分析的其他PaLCuCNV各分离物核苷酸相似性达到91%以上(图4)。依据目前国际双生病毒分类标准,侵染广西辣椒的双生病毒分别属于TYLCV和PaLCuCNV的分离物,各序列在GenBank的登录号为:BS66-1(GenBank登录号:OQ190458)、BS67-1(GenBank登录号:OQ383543)、BS66-2(GenBank登录号:OQ383309)。
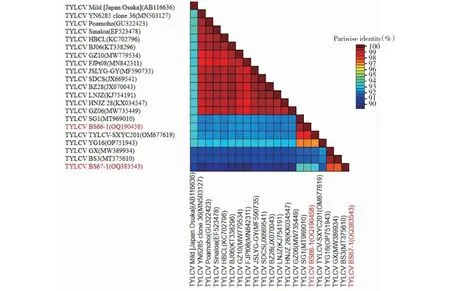
图3 TYLCV广西辣椒分离物BS66-1、BS67-1与其他19个TYLCV分离物相似性比对Fig.3 Sequence identity comparison of TYLCV Guangxi pepper BS66-1,BS67-1 isolates and other 19 isolates of TYLCV
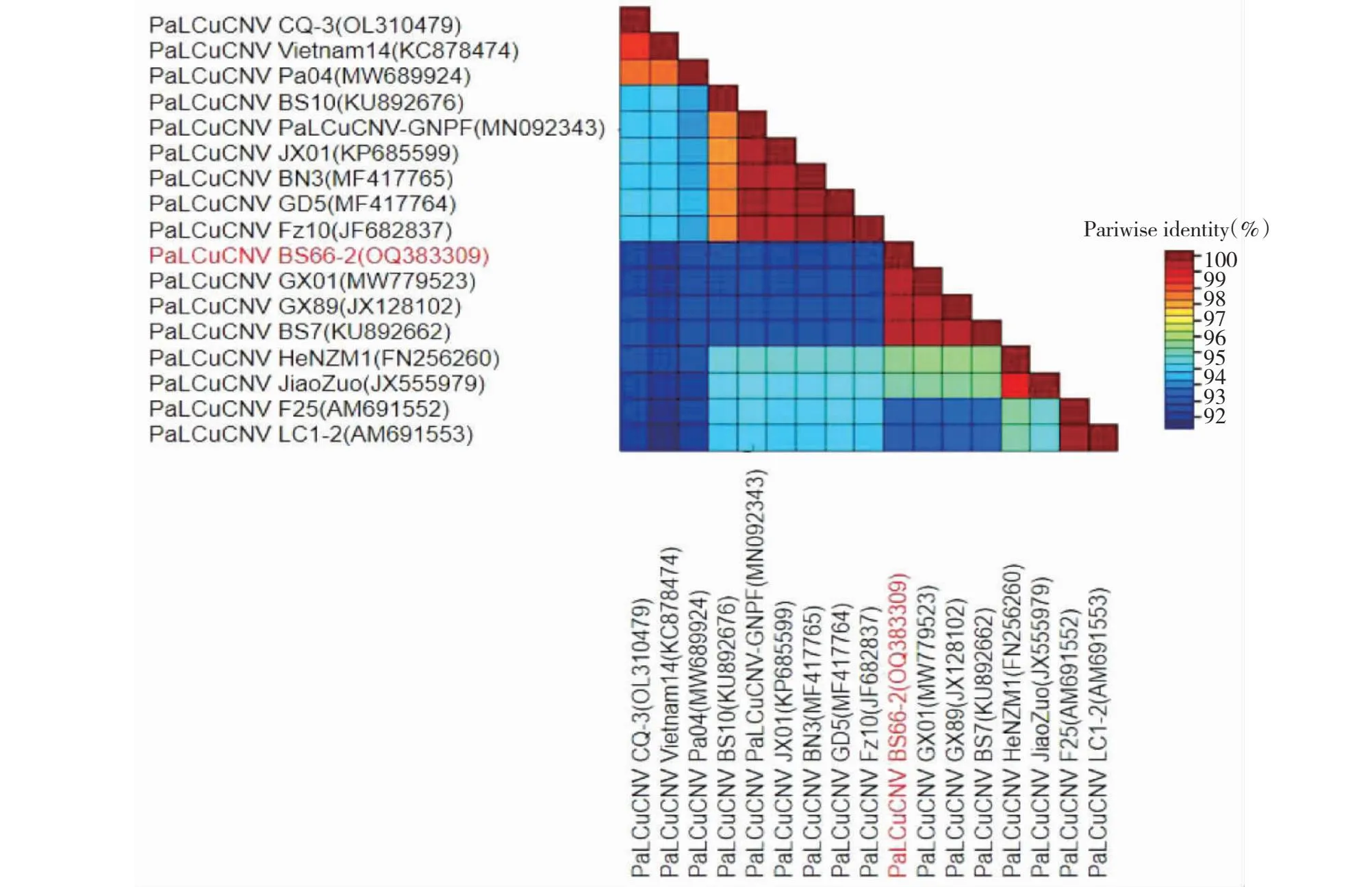
图4 PaLCuCNV广西辣椒分离物BS66-2与其他16个PaLCuCNV分离物相似性比对Fig.4 Sequence identity comparison of PaLCuCNV Guangxi pepper BS66-2 isolates and other 16 isolates of PaLCuCNV
2.3 TYLCV广西辣椒分离物的遗传进化分析
为了解本研究中得到的2个TYLCV广西辣椒分离物与已登录Genkbank的19个TYLCV分离物的进化关系,利用MEGA 7.0软件进行进化树的构建,最大似然法构建进化树为最优树(图5)。本研究获得的2个序列处于不同分支,其中BS66-1与TYLCV中国SG1分离物(GenBank登录号:MT969010,寄主:番茄)处于同一个小分支,说明两者具有较近的亲缘关系;而BS67-1则与中国广西的GX分离物(GenBank登录号:MW389934,寄主:番茄)和BS3分离物(GenBank登录号:MT375610,寄主:番茄)处于相同分支,说明三者具有较近的亲缘关系,但BS67-1又处于1个独立的分支,可能又具有相对独立的进化。
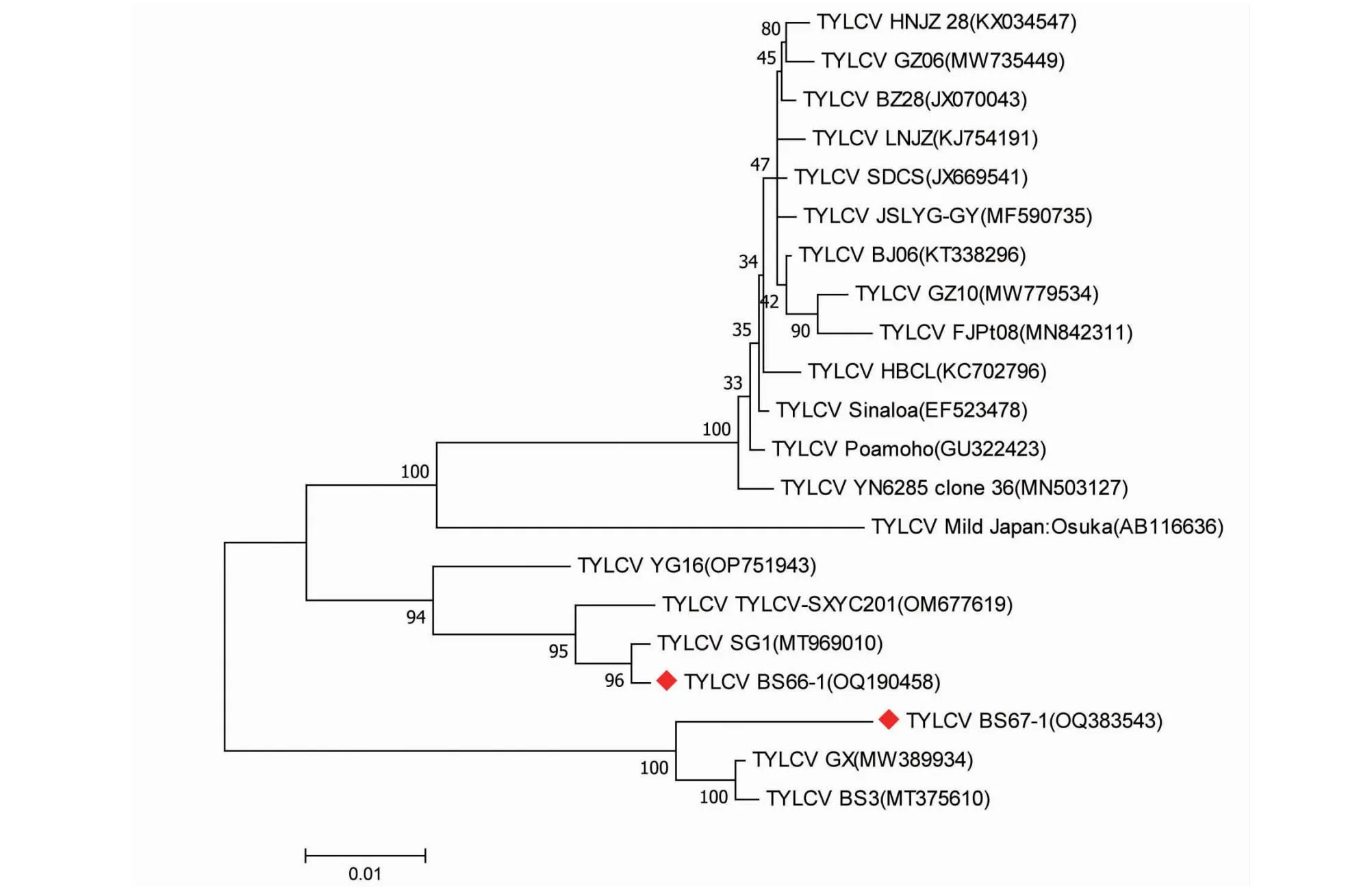
图5 TYLCV广西辣椒分离物BS66-1、BS67-1与其他19个分离物的全长核苷酸序列构建的系统发育进化树Fig.5 Phylogenetic tree of TYLCV Guangxi pepper BS66-1,BS67-1 and other 19 isolates based on full-length sequence
2.4 PaLCuCNV 广西辣椒分离物的遗传进化分析
将PaLCuCNV广西辣椒分离物和已登录GenBank的16个PaLCuCNV分离物用MEGA 7.0软件中的最大似然法进行进化树构建(图6),了解本研究中得到的1个PaLCuCNV广西辣椒分离物与已报道的PaLCuCNV分离物的进化关系。PaLCuCNV广西辣椒分离物BS66-2与PaLCuCNV中国广西3个分离物处于同一个大分支,并且PaLCuCNV广西辣椒分离物BS66-2与PaLCuCNV GX01(GenBank登录号:MW779523,寄主:番茄)处于同一个小分支,说明三者亲缘关系较近,但广西辣椒分离物与PaLCuCNV GX01具有更近的亲缘关系。
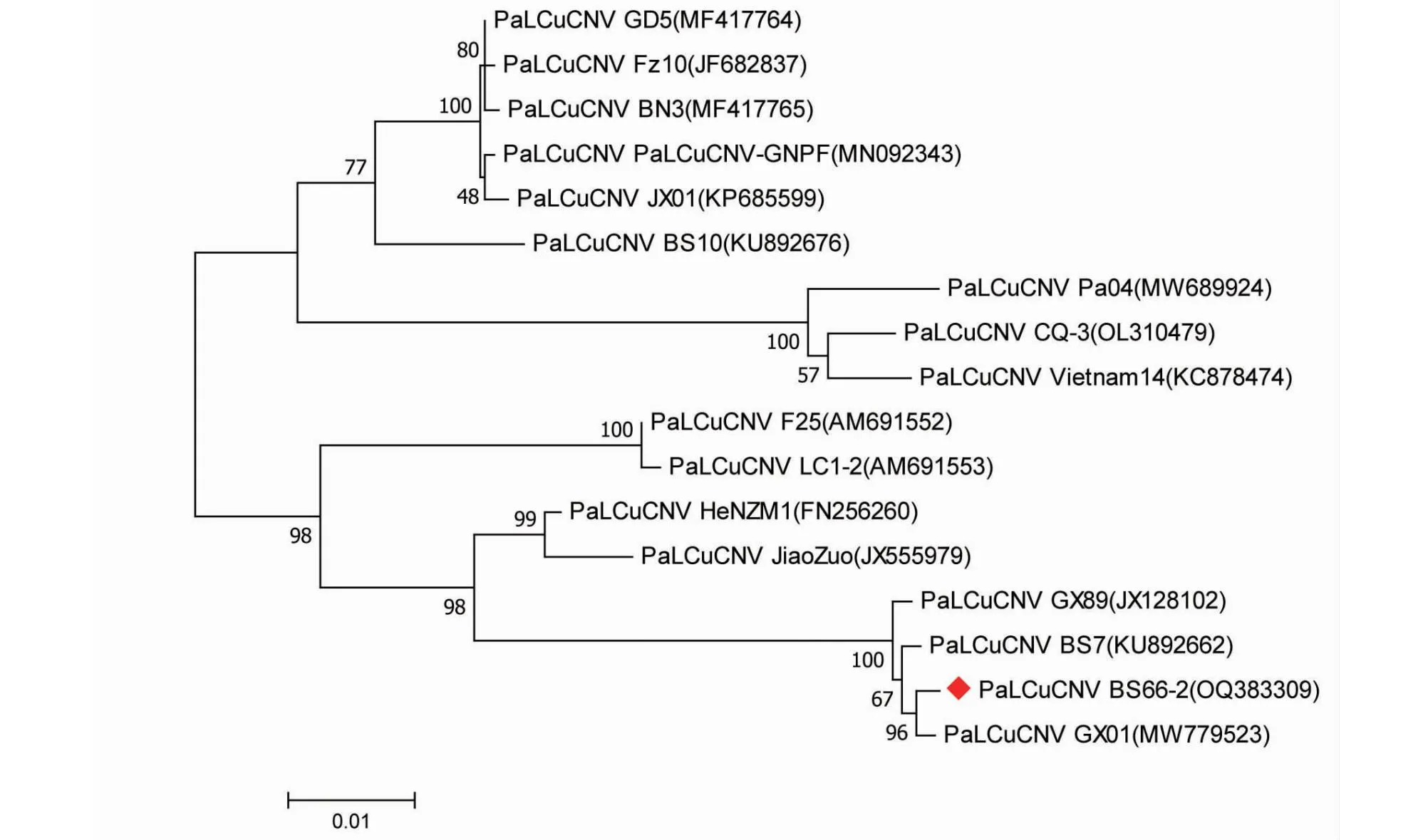
图6 PaLCuCNV广西辣椒分离物BS66-2与其他16个分离物的全长核苷酸序列构建的系统发育进化树Fig.6 Phylogenetic tree of PaLCuCNV Guangxi pepper BS66-2 and other 16 isolates based on full-length sequence
3 讨 论
本研究对广西百色市叶片表现出黄化、皱缩、叶片上卷等症状的辣椒进行鉴定后证实,辣椒受到双生病毒的侵染,进一步利用分段克隆、核苷酸序列比对分析、进化树构建等方法,从2个辣椒阳性样品中共获得3条双生病毒的全基因序列,这3条序列分别与番茄黄化曲叶病毒不同分离物的序列和中国番木瓜曲叶病毒不同分离物序列的核苷酸相似性均在91%以上,根据国际病毒分类委员会对菜豆金色黄花叶病毒的分类标准[20],病毒DNA-A的核苷酸相似性大于91%则认为是同种病毒的不同分离物。因此,侵染广西辣椒的病毒分别属于TYLCV和PaLCuCNV分离物。本研究是TYLCV和PaLCuCNV侵染广西辣椒的首次报道。
通过构建进化树的方式对本研究中获得的3条序列进行遗传进化分析发现,本研究获得的TYLCV BS66-1与TYLCV中国分离物SG1(GenBank登录号:MT969010)处于同一个小分支,说明其亲缘关系较近;而TYLCV BS67-1则与TYLCV中国广西分离物GX(GenBank登录号:MW389934)处于同一个小分支,说明其亲缘关系较近。PaLCuCNVBS66-2则与PaLCuCNV中国河南、广西分离物处于相同大分支,亲缘关系较近。
TYLCV和PaLCuCNV均属于双生病毒科的菜豆金色花叶病毒属病毒,TYLCV最早于1934—1946年在以色列被发现,1964年被正式命名[21-22]。在中国,TYLCV自2006年在浙江、上海等地发现以来,快速蔓延至山东、河南、北京、广东等地[23-24]。TYLCV于2019年传入广西,在番茄上得到鉴定[25],2年后又被再次确认侵染广西的辣椒,证实该病毒正在广西区内扩散蔓延。PaLCuCNV以番木瓜为自然寄主,首次在广西被Wang等[26]发现,主要分布在云南、广西、广州等地[27-29],本研究则证实辣椒是PaLCuCNV的自然寄主。
广西是我国重要的冬种蔬菜生产基地,承担着南菜北运的重要使命。辣椒是广西蔬菜生产中重要的品种,辣椒产业也是广西部分地区收入增长的主要来源[30]。本研究在百色市田阳区的调查中发现,番茄与辣椒混合种植较为普遍,栽种模式增加了不同作物间病毒互相传染的概率。而田阳种植的番茄种苗大多为本地生产,但也存在从福建、云南、四川等多地调苗的现象,番茄种苗区外调运的现象则加剧了区域间虫媒病毒的传播,适宜的传播环境更易造成病毒病害的扩散蔓延。因此,应加强检疫,防止病毒通过种苗传播蔓延,另外,也应加强对该类病毒的监测,及时了解病毒种类动态变化。同时,生产中应种植抗病品种,控制烟粉虱的数量,以此达到控制病毒传播,防范病毒蔓延的目的。
4 结 论
广西百色市辣椒呈现叶片上卷、黄化、皱缩等症状的病毒病原为TYLCV和PaLCuCNV。通过进化树分析发现,本研究获得的TYLCV和PaLCuCNV辣椒分离物与GenBank已报道的TYLCV和PaLCuCNV番茄分离物均具有较近的亲缘关系,说明这2种病毒可能是经番茄传播至辣椒,今后应加强监测。
