干旱胁迫对马缨杜鹃生长和光合生理特征的影响
2023-03-13李苇洁林世成王洪飞周玉梅
曾 松,欧 静,田 奥,李苇洁,林世成,王洪飞,周玉梅
(1.贵州大学林学院,贵阳 550025;2.贵州省山地资源研究所,贵阳 550001)
【研究意义】杜鹃属(Rhododendron)植物是重要的观赏花卉,特别是常绿杜鹃亚属杜鹃,其花大色艳,具备广阔的开发前景[1]。常绿杜鹃亚属杜鹃主要集中分布于我国西南地区光线较强、水分较少的林缘和小山丘等环境[2],大多为大灌木或小乔木,且其根系浅薄,易受环境因子影响[3]。近年来,西南地区持续增温,降水逐年减少,干旱已逐渐威胁杜鹃健康生长及分布[4]。通常植物处于干旱环境时,光合作用(气孔导度、光合速率和叶绿素含量等)率先受到影响,其指标发生改变进而影响物质和能源供给。在光合作用影响下,植物开始通过降低株高、地径、叶面积和增大根冠比等改变形态的方式来减少水分散失,提高植株对土壤水分的吸收[5]。无论是生长还是生理指标均有可能因对土壤水分的适应性不同而表现出不同的数量关系和阈值。百里杜鹃是我国重要的杜鹃种质资源库,常绿杜鹃亚属物种超过70%,马缨杜鹃(Rhododendrondelavayi)是其重要的优势种和建群种[6]。近20年来,贵州省发生极端高温、干旱事件的概率达70%[7],马缨杜鹃出现不同程度的退化甚至死亡[8]。因此,研究干旱胁迫对马缨杜鹃生长和光合生理特征的影响,对保护马缨杜鹃资源具有重要意义。【前人研究进展】目前,大量学者关于干旱胁迫对杜鹃影响的研究已涵盖个体、细胞和分子水平[2,9-10],其从个体生长、细胞器结构和细胞水分方面阐述了杜鹃花在不同干旱程度下的适应策略。一些植物生长的水分临界值研究以植物叶片光合生理参数(光合速率、蒸腾速率水分利用效率等)为衡量标准,通过光合生理参数响应水分梯度的曲线计算积分平均值,作为划分植物水分利用情况的依据[11-12]。尽管光合作用可通过一系列复杂过程为植物体提供能源物质[13],综合体现植物应对干旱胁迫的适应机制。但瞬时光合生理参数往往易受气候条件影响[14],不能完全真实地反应植物的实际生长状况。因此,应建立不同指标的数量关系,将生长及生理指标的水分适应范围加以区分,并寻找关键阈值,更好地探究杜鹃在干旱条件下的生长和生理适应机制。【本研究切入点】关于干旱胁迫对杜鹃影响的研究目前尚未能体现各类指标随定量土壤含水量变化而变化的趋势,不利于关键阈值的确定。【拟解决的关键问题】以2年生马缨杜鹃实生苗为研究对象,采用盆栽控水法结合单因素方差分析和回归分析研究不同干旱胁迫对马缨杜鹃幼苗生长和光合生理指标的影响,建立不同指标响应水分梯度的数量关系,探明适宜马缨杜鹃生长的土壤含水量阈值,揭示其抗旱能力和适应机制,为马缨杜鹃资源的开发利用和抗旱新品种的选育提供依据。
1 材料与方法
1.1 试验材料
1.1.1 杜鹃 杜鹃苗,2年生马缨杜鹃实生幼苗,无病虫害,长势一致,购于贵州省毕节市百里杜鹃万绿源种苗公司。
1.1.2 土壤基质 由黄壤土与腐殖土按质量比1∶4配成,自制。
1.1.3 仪器设备 TR-8D土壤水分仪,顺科达科技有限公司;UV1901紫外分光光度计,上海奥析科学仪器有限公司;Li-6400系列便携式光合作用测量系统, 美国LI-COR公司生产。
1.1.4 其他 塑料盆,上口径17 cm,下口径15 cm,高17 cm。
1.2 试验设计
1.2.1 材料预处理 2021年12月15日,将马缨杜鹃幼苗移栽于装有2 kg土壤基质的塑料盆中,缓苗2个月备用,其间进行正常水分管理。
1.2.2 试验设计 试验共设6个处理。对照(CK):土壤含水量为田间持水量80%~100%;FC60:土壤含水量为田间持水量60%~80%;FC40:土壤含水量为田间持水量40%~60%;FC30:土壤含水量为田间持水量30%~40%;FC20,土壤含水量为田间持水量20%~30%;FC10:土壤含水量为田间持水量10%~20%。每个处理栽20盆,每盆栽1株。2022年2月15日,充分浇水使马缨杜鹃盆栽土壤含水量饱和,并使其自然落干,随后每天用土壤水分仪检测盆栽土壤含水量,当达到各预设土壤干旱程度时记录盆栽重量,并按处理进行分组。至2022年3月1—5日,试验设定的各处理和重复均已达到要求,自此开始干旱胁迫试验,此后每天18:00称重控水,及时补足盆土散失的水分。为减少马缨杜鹃自身生长重量的影响,每隔5 d用土壤水分仪检测土壤含水量,及时调整浇水量。连续控水3个月后,于6月15日测定不同干旱处理下马缨杜鹃的形态指标和光合生理指标。
1.2.3 指标测定 (1)形态指标。采用卷尺测定各处理马缨杜鹃的株高(栽培基质表面至马缨杜鹃幼苗主茎顶端的距离),采用游标卡尺测定地径,每株地径为水平面垂直方向测2次取平均值;使用卷尺测定芽长(芽痕至芽先端长度),每个处理测定20株主干顶芽长取平均值;通过计算株高和地径的相对生长量HO(%)及DO(%)进行比较。叶面积采用网格法测定,每个处理随机选择5株马缨杜鹃幼苗,分别取当年生第3轮新叶和上一年生第3轮老叶紧贴于0.5 cm×0.5 cm的网格纸上,用铅笔描出轮廓,数出新叶和老叶覆盖的方格数,分别计算新叶和老叶的面积(不足半格的舍去,超过半格的计1格)。采用目数法测定干旱胁迫开始时每盆植株的老叶数及胁迫试验之后每盆植株的新叶数和落叶数,并比较干旱胁迫处理后植株的新叶率(RN,%)、落叶率(RO,%)及留在植株上的总叶面积(AT,cm2)。叶片相对含水量(RWC,%)采用烘干法测定,从每个处理随机选择5株马缨杜鹃幼苗,取第3轮当年生成熟叶片带回实验室测定相对含水量。
(1)
(2)
(3)
(4)
AT=N×AN+O×AO-F×AO
(5)
(6)
式中,HO(%)及DO(%)分别为株高和地径的相对生长量,H1(cm)、D1(mm)及H2(cm)、D2(mm)分别为初始和干旱胁迫3个月后的株高、地径,RN(%)及RO(%)分别为新叶率和落叶率,N、O及F分别为新叶数、老叶数及落叶数,AT(cm2)、AN(cm2)和AO(cm2)分别为总叶面积、新叶面积和老叶面积,M1、M2和M3分别为叶片鲜重、干重和吸水饱和重。
(2)光合生理指标。叶绿素采用乙醇浸提法测定[15],即从每个处理随机选择5株马缨杜鹃幼苗,每株取1张第3轮叶片用蒸馏水洗净叶子表面杂质并擦干水分,去掉中脉剪碎混匀后,准确称取0.1 g马缨杜鹃叶子剪碎放入离心管中,加入8 mL 95%乙醇,浸提24 h充分提取光合色素。提取完毕后,吸取1 mL浸提液加入比色皿,再加入2 mL 95%乙醇稀释,在波长分别为470和645 nm的紫外分光光度计下测定吸光值,再计算叶绿素a和叶绿素b的含量。光合参数:采用Li-6400便携式光合作用测定系统进行测定,即选取6月中旬晴朗无风天气,在9:30—12:00进行叶片各项光合参数的测定。开始测定前,利用光合仪自带红蓝光叶室,将光源有效辐射强度设置为1000 μmol/(m2·s),选取第3轮成熟叶片活化30 min,待各项指标稳定后开始测定。
(3)土壤水分缓冲区的确定。随着土壤含水量的逐渐降低,生长和生理指标表现为2种响应形式:一种为单调递增或递减变化,一种为单峰曲线变化。对于呈单调变化的指标,土壤水分梯度直接反映指标变化水平,没有在轻微干旱的时候产生适应性而升高。对于呈单峰曲线变化的指标,当土壤含水量由CK逐渐降低,相应指标表现为先缓慢增加至最大值后逐渐下降至CK对应的初始值水平,认为在该区间内的指标对轻微的干旱产生一定的适应性而适当增加,之后随土壤含水量的降低继续下降,将CK对应的指标初始值水平之间的土壤含水量范围定义为土壤水分缓冲区。
1.3 数据统计与分析
采用Excel 2010对不同干旱程度下马缨杜鹃的生长和光合生理指标进行记录,使用SPSS 19.0进行单因素方差分析和Duncan多重比较(P<0.05)。采用OriginPro 2016进行回归分析并绘制图形。
2 结果与分析
2.1 干旱胁迫处理马缨杜鹃的株高和地径
从图1看出,不同处理马缨杜鹃在3月15日的初始株高基本一致,均保持在20 cm左右,而初始地径除FC60为6.08 mm外,其他处理初始地径均在5.50 mm左右。干旱胁迫3个月后,马缨杜鹃株高为26.18~39.11 cm,随干旱胁迫加剧呈先降后升再降趋势,虽FC30大于FC40,但各处理均低于CK,整体还是呈随干旱加剧逐渐减小趋势;地径为5.84~6.81 cm,随干旱加剧呈先升后降趋势,FC60具有最大值。株高相对生长量为25.68%~49.86%,随干旱胁迫加剧呈先升后降再升再降趋势,FC60显著高于FC40、FC20和FC10,与其余处理差异不显著;FC40显著低于FC60和FC30,FC10显著低于其余处理。地径相对生长量为8.92%~13.03%,各处理间差异不显著。说明,重度干旱显著限制马缨杜鹃株高生长。通过回归分析可知,马缨杜鹃株高和地径相对生长量随土壤含水量增加均呈先增后减的响应趋势。其中,株高相对生长量与土壤含水量拟合精度(R2=0.67,P=0.19)高于地径相对生长量(R2=0.37,P=0.48),但二者均未达显著水平。
2.2 干旱胁迫处理马缨杜鹃叶片的生长
由表1可知,不同干旱胁迫马缨杜鹃叶片的生长存在明显差异。芽长为0.63~4.60 cm,随干旱胁迫加剧呈先升后降趋势,其中,FC60显著高于FC10、FC20和FC30,FC10显著低于除FC20外的其余处理,其余处理差异显著或不显著。新叶数为8.82~21.86片,随干旱胁迫加剧呈先升后降趋势,其中,FC60显著高于FC30、FC20和FC10,其余处理差异不显著。落叶数为2.00~4.36片,随干旱胁迫加剧呈先升后降再升趋势,其中,FC20、FC10显著大于CK,其余处理间差异不显著。总叶数为21.36~34.36片,随干旱胁迫加剧呈先升后降趋势,其中,FC60显著高于FC10,其余处理间差异不显著。单叶面积为25.33~70.42 cm2,随干旱胁迫加剧呈下降趋势,其中,CK显著高于其余处理,FC60显著高于除FC40外的其余处理,FC40显著高于FC20和FC10,FC30与FC20之间差异不显著但均显著高于FC10。叶片相对含水量为0.74%~0.99%,仅FC10显著低于其他处理。综合看,不同处理叶片的生长指标对干旱胁迫的响应不尽一致,总体上过低的土壤水分含量会显著抑制叶片的生长。

柱状图上不同小写字母表示不同处理间差异显著(P<0.05)。Different lowercase letters indicate significance of difference among different treatments at P<0.05 level.
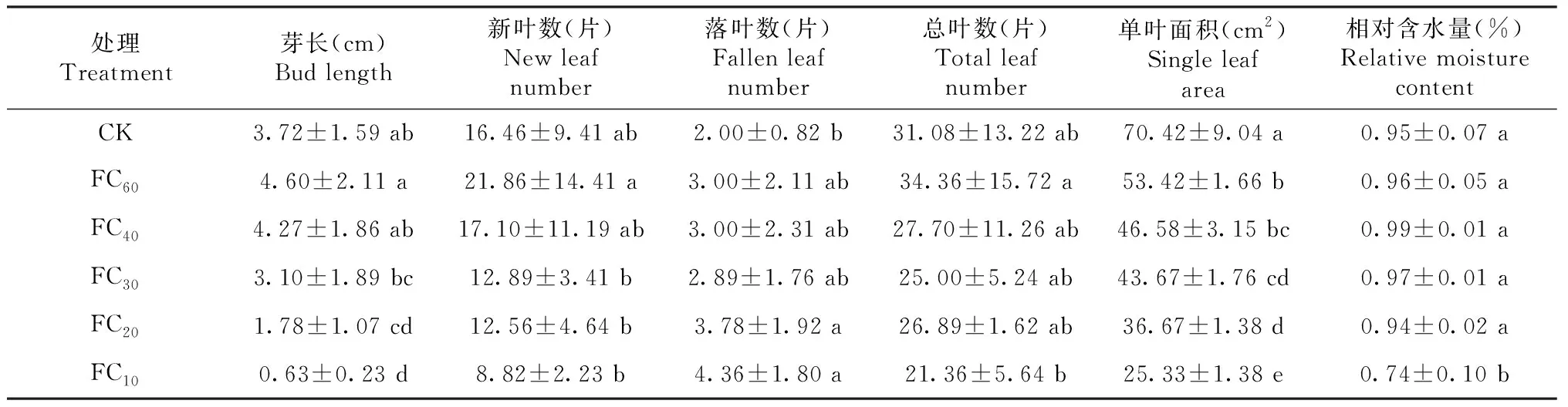
表1 不同干旱胁迫马缨杜鹃叶片的生长
由图2可知,除叶片相对含水量与土壤含水量的回归效果未达显著水平(R2=0.77,P=0.11)外,其余叶片生长指标均与土壤含水量存在显著回归关系。其中,落叶率随土壤含水量增加呈逐渐降低趋势,总叶面积和单叶面积随土壤含水量增加均呈逐渐增加趋势,表明,马缨杜鹃通过减少叶面积,促进老叶掉落的方式适应干旱胁迫。芽长、新叶率及叶片相对含水量随土壤含水量增加呈先升后降的单峰曲线变化,其中,芽长在土壤含水量为32.78%(CK)时为3.7 cm,随着土壤含水量逐渐降低至25.97%,芽长缓慢增加到最大值4.61 cm,此后土壤含水量继续降低至16.30%时,芽长开始逐渐减少至初始CK处理下的3.7 cm,此后随干旱胁迫的持续加剧,芽长开始迅速下降。由此可知,土壤含水量为25.97%时是马缨杜鹃芽长的最适土壤含水量,而土壤含水量介于16.30%~32.78%时,芽长生长并未在干旱胁迫下减少,反而产生了一定的适应性,因此土壤含水量为16.3%~32.78%是芽长在干旱胁迫时的缓冲区。同样,马缨杜鹃新叶率达最大值53.63%的最适土壤含水量为25.18%,土壤干旱胁迫缓冲区阈值为15.08%~32.78%。

图2 叶片生长指标与土壤含水量的回归曲线模拟Fig.2 Regression curve simulation of leaf growth index and soil moisture content
2.3 干旱胁迫处理马缨杜鹃的光合特性
由表2可知,不同干旱胁迫马缨杜鹃叶片的光合特性存在明显差异。
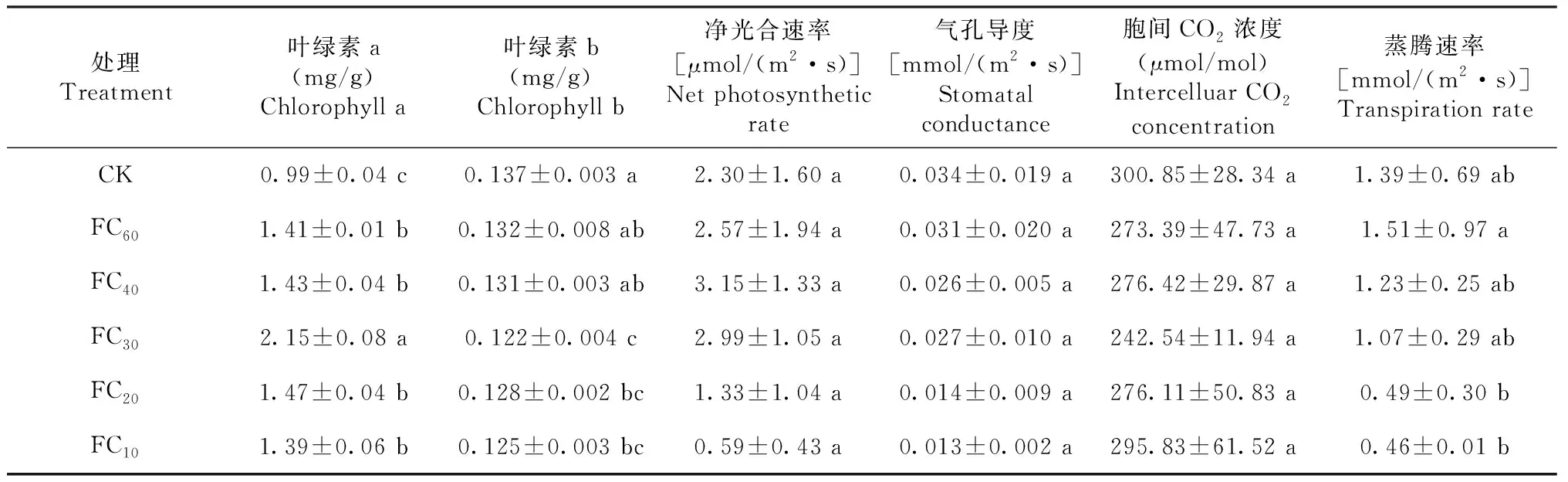
表2 不同干旱胁迫马缨杜鹃的光合特性
叶绿素a为0.99~2.15 mg/g,随干旱胁迫加剧呈先升后降趋势,其中,CK叶绿素a显著低于其他处理,而FC30显著高于其他处理。表明,干旱胁迫对马缨杜鹃叶绿素a含量具有显著影响。叶绿素b为0.12~0.14 mg/g,随干旱胁迫加剧呈先降后升再降趋势,其中,FC30、FC20和FC10差异不显著但均显著低于CK和FC60、FC40,后三者差异不显著。净光合速率、气孔导度和胞间CO2浓度分别为0.59~3.15 μmol/mol,0.01~0.03 mmol/mol,242.54~300.85 μmol/mol,依次呈先升后降、下降和先降后升趋势,但各处理间差异不显著。蒸腾速率为0.46~1.51 mmol/(m2·s),呈先升后降趋势,其中,FC60显著高于FC20和FC10,其余处理间差异不显著。说明,马缨杜鹃叶绿素对干旱胁迫的响应较敏感,而光合参数中仅蒸腾速率受干旱影响显著。
由图3可知,叶绿素a(P=0.29)及净光合速率(P=0.06)随土壤含水量增加呈先升后降的单峰曲线变化,叶绿素b(P=0.10)随土壤含水量增加呈逐渐增加趋势,胞间CO2浓度(P=0.24)则随土壤含水量的增加呈先降后升趋势,然而这些回归关系均未达显著性水平。气孔导度(P=0.03)及蒸腾速率(P=0.02)随土壤含水量的变化回归效果达显著水平。其中,气孔导度随土壤含水量增加逐渐增加,且增大幅度逐渐平缓;蒸腾速率随土壤含水量增加呈先增后减趋势,在土壤含水量阈值为30.09%时具有最大值1.46 mmol/(m2·s),土壤干旱胁迫缓冲区阈值为27.40%~32.78%。
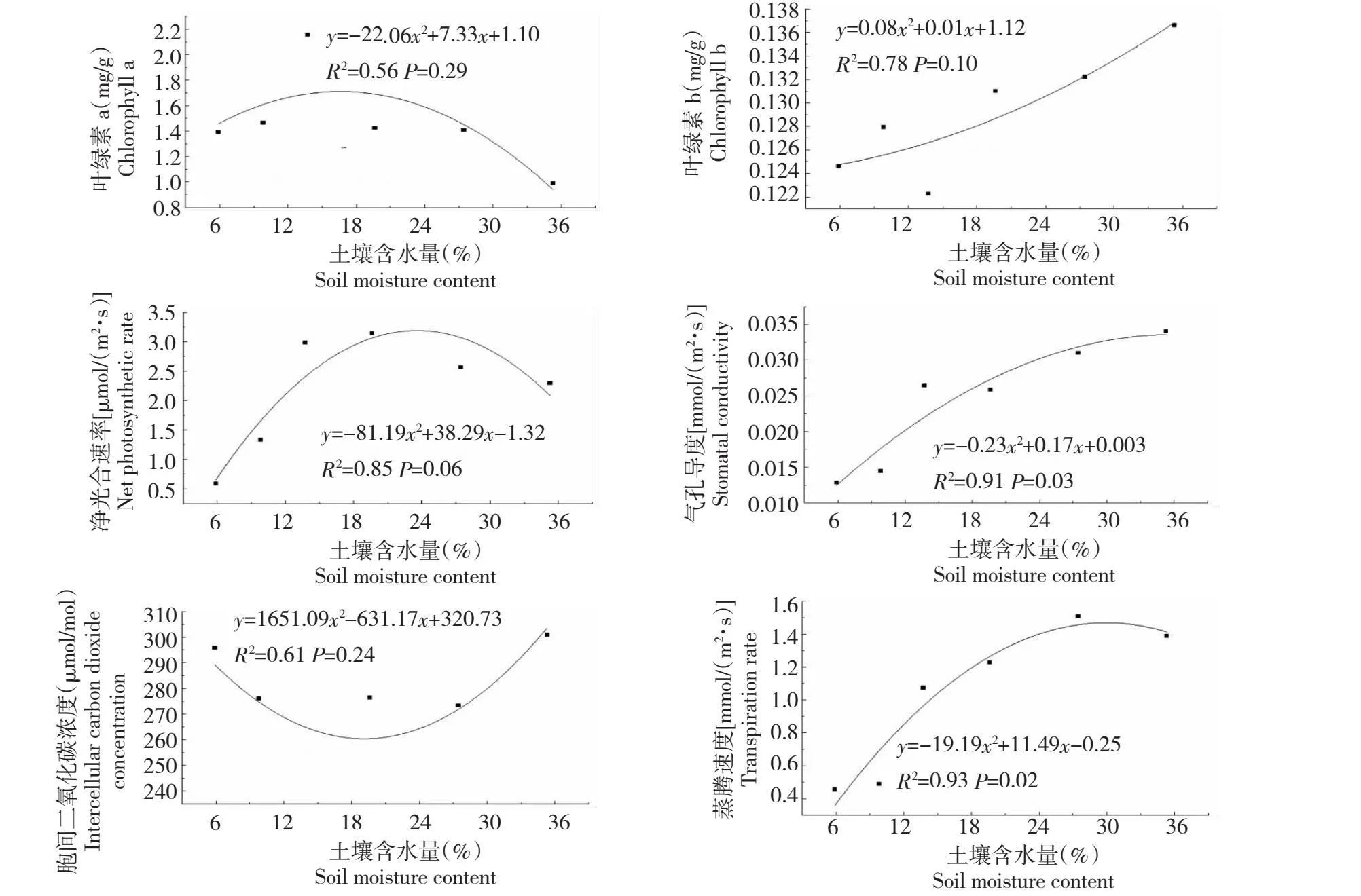
图3 光合特性与土壤含水量的回归曲线模拟Fig.3 Regression curve simulation of photosynthetic characteristics and soil moisture content
3 讨 论
在应对干旱胁迫时,通过改变形态特征适应土壤水分变化是植物的主要生存策略[16]。有研究表明,黑果腺肋花楸幼苗地径在干旱胁迫下显著降低,而株高变化不显著[17]。本研究结果表明,不同干旱胁迫马缨杜鹃的株高增长量存在明显差异,且随着干旱胁迫程度降低,株高相对生长量逐渐增加,当土壤含水量为26.00%时达最大值,之后开始缓慢降低,尽管回归关系并未达显著水平,但拟合精度仍然在可接受范围内。表明,马缨杜鹃对土壤水分有一定的适应范围,土壤水分过高或过低均会限制株高的增长,与苏春莉等[18]的研究结论一致。地径相对生长量在不同干旱胁迫下差异不显著,且其回归关系也不显著,拟合精度较低,表明,地径生长受干旱影响较小,与张芸香等[19]研究杜仲地径对干旱胁迫响应的结论一致。这是因为植物主干是水分运输通道,受土壤水势的影响明显,即干旱条件下土壤水势明显降低,水分运输距离缩短,株高生长明显受到影响,因此株高对土壤干旱的响应更为敏感,马缨杜鹃主要通过降低株高来减少土壤水分的消耗,以适应干旱[20]。
相较于株高和地径,叶片是植物直接与外界环境进行物质和能量交换的重要器官,对环境变化的响应更加敏感[21]。通过多重比较可知,不同干旱胁迫叶片的生长指标均差异显著。单叶面积和总叶面积随干旱胁迫减弱逐渐增加,落叶率随干旱胁迫增强逐渐增大,与丁雪丹等[22]的研究结论一致。然而,芽长、新叶率和叶片相对含水量并未表现出随干旱胁迫增强而单调递减的变化趋势,而是先缓慢增加后逐渐降低,反映出叶片生长在抵御轻微干旱时,产生了一定的适应性;其相应土壤含水量的缓冲区域分别为16.30%~32.78%、15.08%~32.78%和12.32%~32.78%。相似的变化结果在王园园等[23]对苜蓿干旱胁迫的研究中也得到印证。一般情况下,较低的土壤水分环境可促使植物通过自我调整叶片数量和面积从而使体内资源利用最大化,以适应胁迫环境[24]。本研究结果表明,在干旱胁迫水平较低时,马缨杜鹃可通过降低叶片面积、增加老叶凋落数量的策略来适当提高芽长及新叶增长数量;当干旱胁迫持续加重,超过土壤含水量缓冲区的阈值后,叶片的各生长指标均迅速降低。
叶绿素a和叶绿素b在植物光合作用过程中参与光能的吸收、耗散和分配[8]。研究结果表明,干旱胁迫下叶绿素a的含量显著高于CK,叶绿素b的含量也在干旱胁迫程度小于FC30时显著降低。叶绿素a和叶绿素b的含量分别反映植物对长波和短波光的吸收能力[24],马缨杜鹃对长波光和短波光的吸收能力分别与青冈栎及茄子一致。尽管净光合速率、气孔导度及胞间CO2浓度在不同干旱胁迫下差异均不显著,但气孔导度对干旱梯度的响应仍然达显著水平。气孔导度反映植物细胞与外界交换气体的能力[25],干旱加剧可使气孔皱缩、通道变窄[26],限制植物蒸腾速率及气体交换能力。然而本研究中气孔导度与蒸腾速率、植物生长指标(株高、芽长、新叶率、叶片相对含水量等)对干旱胁迫的响应并不一致,表明,在轻度干旱条件下,植物光合速率下降并非受气孔的限制[17,25],而是由于叶肉细胞光合活性下降或根系吸水能力降低所致。
4 结 论
不同干旱胁迫对马缨杜鹃幼苗生长及光合特征的影响存在明显差异。其中,马缨杜鹃幼苗株高增量在最高和最低土壤含水量下差异显著;叶片生长指标中,FC60和FC10的叶长、新叶数、总叶数和单叶面积相对含水量具有显著差异,芽长、新叶率、落叶率、总叶面积及单叶面积与土壤含水量的回归关系显著。其中,芽长和新叶率在土壤含水量分别为25.97%和25.18%时达最大值;土壤含水量分别为16.30%~32.78%和15.08%~32.78%是马缨杜鹃幼苗适应干旱胁迫的缓冲区。光合特征指标中仅叶绿素a、叶绿素b和蒸腾速率在不同干旱胁迫下差异显著,仅气孔导度及蒸腾作用与土壤含水量的回归关系达显著水平。其中,蒸腾作用在土壤含水量为30.09%时达最大值,土壤含水量为27.40%~32.78%时为马缨杜鹃幼苗适应干旱胁迫的缓冲区。
