母乳低聚糖在婴幼儿生长发育中的作用及其影响因素
2023-03-07蒋思瑾
蒋思瑾,石 璐,周 静,杨 静,程 悦,韩 蓓
(1.西安交通大学医学部公共卫生学院,陕西 西安 710061;2.西安交通大学第二附属医院儿科,陕西 西安 710004;3.西安市临潼区妇幼保健院营养科,陕西 西安 710699)
母乳是婴幼儿最天然、最营养的食物来源,对婴幼儿的生长和发育至关重要。世界卫生组织和联合国儿童基金会建议,婴儿出生后应尽早开始母乳喂养,6个月内纯母乳喂养,之后配合辅食持续母乳喂养至2岁或更长[1]。母乳成分是母乳喂养的主要优势之一,除了含有常规的营养成分(蛋白质、脂肪、碳水化合物、维生素、矿物质、水),母乳中还含有大量特殊的生物活性成分,如母乳低聚糖(human milk oligosaccharides,HMOs)、活性蛋白、生长因子、细胞因子、微生物等[2]。HMOs虽然不能被婴幼儿吸收代谢,但能被肠道微生物利用,并选择性刺激双歧杆菌、乳杆菌等肠道有益菌的生长,对肠道菌群稳定和肠道免疫屏障的形成具有积极的影响[3]。越来越多的研究证明母乳低聚糖对婴幼儿健康有着重要作用,因此本文对当前国内外母乳低聚糖的重要作用及其影响因素的研究进行梳理,现综述如下。
1 母乳低聚糖的种类和结构
HMOs是母乳中第三丰富的固体成分,仅次于乳糖和脂肪,约占母乳碳水化合物的20%,HMOs是人类特有的,其他哺乳动物中没有相同的低聚糖分子和组成。牛乳成分的婴儿配方奶粉中缺乏HMOs,是导致配方奶粉喂养婴儿免疫紊乱发病率高的主要原因[4]。HMOs可分为三类:中性岩藻糖基低聚糖、中性非岩藻糖基低聚糖和唾液酸化低聚糖,目前已经被确定结构的HMOs有200多种[2]。HMOs主要由5种单糖构成,分别是D-葡萄糖(Glucose,Glc)、D-半乳糖(Galactose,Gal)、N-乙酰葡萄糖胺(N-acetylglucosamine,GlcNAc)、L-岩藻糖(Fucose,Fuc)和N-乙酰神经氨酸(唾液酸,Sialic acid,Neu5Ac)[4]。HMOs能够抵抗胃肠道的水解和消化,且无法被小肠吸收,因此对宿主而言它们几乎没有营养功能,但能被肠道微生物代谢,产生各种有益作用[3]。
2 HMOs与婴儿健康
2.1 HMOs促进肠道内有益菌增殖,调节婴儿肠道菌群平衡
婴儿肠道菌群与婴儿的发育和免疫系统的形成密切相关[5]。母乳喂养的婴幼儿,随着母乳消化吸收而到达婴儿肠道的HMOs,一部分随粪便排出体外,另一部分在婴儿肠道中被某些有益菌作为底物发酵产生短链脂肪酸(short chain fatty acids,SCFAs)和乳酸,从而创造了一种不利于大多数病原微生物生长的酸性环境[6]。HMOs能够选择性地促进肠道中的有益细菌生长,尤其是某些双歧杆菌、乳酸杆菌和拟杆菌,形成健康的肠道微生物群[7]。研究表明,母乳喂养婴儿的粪便细菌组成与婴儿配方乳粉喂养的婴儿不同;虽然双歧杆菌在母乳喂养婴儿和配方乳粉喂养婴儿的粪便中均为主导菌群,但不含HMOs的婴儿配方乳粉喂养的婴儿粪便中双歧杆菌含量低于母乳喂养的婴儿;除双歧杆菌外,母乳喂养婴儿的粪便中乳酸杆菌和链球菌较多,而婴儿配方乳粉喂养婴儿的粪便中葡萄球菌、大肠埃希菌和梭状芽孢杆菌较多[8]。
2.2 HMOs保护婴儿免受病原体入侵
大量体内体外研究表明,HMOs及其代谢产物具有拮抗病原体入侵的作用。由于HMOs和肠黏膜细胞上与微生物结合的多糖含有类似的抗原表位,可以作为可溶性诱饵受体竞争性地结合病原体,减少其与肠黏膜结合的机会,因此HMOs在肠道的存在可以有效阻止病原体的附着和入侵肠上皮[8],见图1。不同结构的HMOs能够结合不同的致病菌,例如2′-岩藻糖基乳糖(2′-fucosyllactose,2′-FL)可阻止空肠弯曲菌与肠黏膜结合,预防空肠弯曲菌感染及其产生的黏膜炎症[2];3′-唾液酸乳糖(3′-sialyllactose,3′-SL)可以抑制肠致病性大肠埃希菌与肠黏膜结合所引起的感染性腹泻和败血症等[7]。
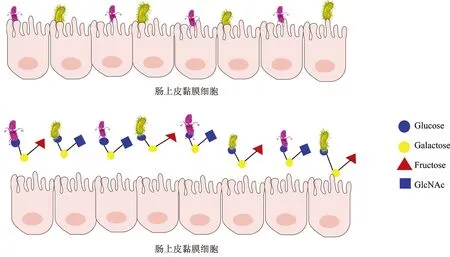
图1 肠道内的HMOs保护婴儿免受病原体入侵的作用机制示意图Fig.1 Schematic diagram of the mechanism of HMOs in the intestine to protect infants from pathogen invasion
HMOs还可以预防病毒对婴幼儿的感染。有研究表明,2′-FL可显著抑制G1P[8]型轮状病毒感染,而3′-SL和6′ -唾液酸乳糖(6′-sialyllactose,6′-SL)对G2P[4]型轮状病毒感染的抑制能力最强[9];组织血型抗原(histo-blood groupantigens,HBGAs)是诺如病毒粘附的关键结合位点和碳水化合物表位,不仅存在于红细胞表面,也存在于胃肠道、泌尿生殖道和呼吸道的黏膜上皮中,由于α-岩藻糖基HMOs与HBGAs的结构相似,因此诺如病毒可通过与α-岩藻糖基HMOs结合[10],而减少其对婴幼儿的感染。
2.3 HMOs促进婴儿大脑发育
唾液酸是支持婴幼儿大脑发育的重要营养物质,婴幼儿需要足够的唾液酸以维持大脑和中枢神经系统的正常发育和功能[7]。成人自身可由肝脏合成内源性唾液酸,但婴幼儿的肝脏和其他器官尚未发育成熟,自身合成的唾液酸并不能满足身体的需要[11]。孕期妇女的唾液和血浆中有高浓度的唾液酸,可通过胎盘进入胎儿体内,保障了胎儿对唾液酸的需求;对于哺乳期的婴幼儿,可以通过母乳获得充足的唾液酸,初乳中唾液酸含量高达1 500mg/L,6个月后降低并维持在250mg/L左右。
有研究表明,参与人脑组织中神经节苷脂和糖蛋白构成的唾液酸大多是以唾液酸化HMOs形式存在,与神经突触和神经传导关系密切,能促进婴儿的认知发育,增强学习和记忆能力[12]。而唾液酸化HMOs是母乳HMOs的3个主要组分之一,特别是在初乳中,占总量的20%~30%。一项分析猝死婴儿的额叶皮层样本的研究发现,接受母乳喂养的婴儿额叶皮质灰质中的神经节苷脂结合和蛋白质结合的唾液酸浓度分别比配方奶喂养婴儿的高32%和22%(P<0.01)[13]。Cho等[14]在一项观察性研究中发现3′-SL与认知水平,尤其是语言功能呈正相关关系。因此可以认为,母乳中的唾液酸化HMOs是婴幼儿大脑认知功能发育不可或缺的生物活性物质。
2.4 HMOs促进早产儿康复
早产儿较足月儿在婴儿期更易患病及死亡。坏死性小肠结肠炎(necrotizing enterocolitis,NEC)是早产儿最常见、最致命的肠道疾病之一,母乳HMOs在防治NEC方面发挥关键作用[3]。在过去30年中,世界范围内的NEC死亡率几乎没有变化,重症病例的死亡率则高达25%[15]。NEC的主要危险因素是早产、病原菌定植和婴儿配方奶粉喂养[16]。目前,NEC最有效的防治策略包括母乳喂养、捐赠母乳喂养及益生菌添加喂养[17]。与配方奶粉喂养的早产儿相比,母乳喂养的早产儿患NEC的风险降低了6~10倍[15]。HMOs对NEC的保护机制可能包括抑制肠上皮细胞的Toll样受体4(TLR4)信号[6]、降低肠道通透性、通过选择性增加有益菌来促进有益的肠道微生物群落结构的建立[18]。在一项病例对照试验中,Autran等[15]发现NEC病例中几乎所有母乳样本中的二唾液酸基-乳糖基-N-四糖(disialyllacto-N-tetraose,DSLNT),是母乳HMOs中的一种唾液酸化HMOs,其浓度均显著低于对照组,由此推测母乳中DSLNT含量是一种潜在的标记物,可用于识别有NEC风险的婴儿,并筛查高危的捐赠母乳。
3 影响母乳HMOs种类和含量的因素
母乳HMOs的种类和含量受多种因素影响,包括遗传、孕期状况、分娩方式、泌乳阶段、遗传、膳食、环境等,详见图2。

图2 影响母乳HMOs种类和含量的因素Fig.2 Factors affecting the type and content of HMOs in breast milk
3.1 泌乳阶段与HMOs
母乳中的HMOs含量和种类与母亲产后的泌乳阶段有着密切联系。一般而言,在泌乳的早期阶段,HMOs的总体含量较高,由初乳(1~7天)中的20~25g/L到成熟乳(15天以后)中的5~20g/L[12]。而单个HMOs的浓度也是根据泌乳阶段的变化而变化。虽然已被发现的母乳HMOs有200多种,但是以表1中的15种HMOs为主,这15种占HMOs总量的75%[19]。整个泌乳期最丰富的中性岩藻糖基HMOs是2′-FL,最丰富的中性非岩藻糖基HMOs是LNT,最丰富的唾液酸化HMOs是6′-SL[20]。研究发现,大多数HMOs在初乳后表现出减少或保持稳定的趋势,而3′-FL在初乳后表现出增加的趋势[21],这也许意味着3′-FL在婴儿生长发育的后期阶段发挥作用。因为母乳成分发生变化,通常是为了满足生长中婴儿的营养和发育的动态需求。

表1 主要HMOs的名称及分类[19]Table 1 Names and classifications of main HMOs
3.2 遗传因素与HMOs
母亲的遗传和生理状态也是影响HMOs组成的主要因素。岩藻糖基HMOs的组成和含量主要由多种岩藻糖基转移酶(fucosyltransferase,FUT)的活性决定,岩藻糖基转移酶主要由分泌基因(Se)和Lewis(Le)基因编码[20]。Se基因编码岩藻糖基转移酶2(fucosyltransferase 2,FUT2),该酶通过α-1,2键将岩藻糖基连接到HMOs的核心结构上;Le基因编码岩藻糖基转移酶3(fucosyltransferase 3,FUT3),该酶通过α-1,3键或α-1,4键将岩藻糖基连接到HMOs的核心结构上[22]。Se基因和Le基因都有显性表达和隐性表达两种。Se基因显性表达时(Se+),FUT2具有活性,相应的α-1,2键岩藻糖HMOs含量多,如2′-FL、LNFP-I,称为分泌型[12];反之,Se基因隐性表达时(Se-),FUT2不具有活性,相应的α-1,2键岩藻糖HMOs含量较少或检测不到,称为“非分泌型”[12]。Le基因显性表达时(Le+),FUT3具有活性,通过FUT3作用的α-1,3键或α-1,4键的岩藻糖HMOs含量多。当Le基因隐性表达时,母乳缺乏这些具有α-1,3键或α-1,4键的岩藻糖,如LNFP-II和3′-FL[23]。FUT2/3失活遗传变异存在于高达30%的人群中,导致Se-/Le-阴性表型,显著影响母乳HMOs总量和浓度[24]。
FUT2/3失活遗传变异主要是Se和Le基因单核苷酸位点多态性(single nucleotide polymorphism,SNP)引起的。一项研究表明Se基因的SNP位点rs601338的G/A多态性是来自美国的母亲控制母乳α-1,2岩藻糖基寡糖水平的关键位点,但该位点的基因型并不影响中国母亲母乳α-1,2岩藻糖寡糖的表达[25],另一个SNP位点rs1047781的A/T多态性则是控制中国母亲ɑ-1,2岩藻糖基寡糖水平的关键位点[26]。
3.3 膳食因素与HMOs
母乳中HMOs的浓度和成分也会受到哺乳期母亲饮食的影响,研究表明哺乳期妇女膳食中宏量营养素摄入量与HMOs浓度有一定关联。Azad等[27]发现总蛋白质摄入量和唾液酸-乳糖-N-四糖b(Sialyllacto-N-tetraose b,LSTb)之间存在微弱的负相关,该研究还发现谷物摄入量和岩藻糖基乳糖-N-六糖(Fucosyllactose-N-hexose,FLNH)之间存在正相关。Quin等[28]的研究中报告,几种磺化的HMOs与单不饱和脂肪和多不饱和脂肪摄入量呈正相关,与饱和脂肪和膳食胆固醇摄入水平呈负相关。近期的研究也证实了改变哺乳期妇女的饮食,特别是碳水化合物的来源,会在短时间内导致一些主要HMOs成分的改变。在一项交叉对照喂养研究中,Seferovic等[29]发现高脂肪饮食(>40%的总能量)导致唾液酸化HMOs浓度降低,葡萄糖与半乳糖饮食相比更容易影响岩藻糖基HMOs的分布。在这项研究中将哺乳期妇女随机分到摄入特定的膳食碳水化合物—葡萄糖组或半乳糖组,经过30~57h严格饮食再经过1~2周“洗脱”期后再进行自身交叉试验,结果表明相对于半乳糖饮食,葡萄糖饮食的母乳中岩藻糖基化HMOs显著减少,而唾液酸化HMOs无明显变化。该研究还将哺乳期妇女随机分配到高碳水化合物饮食或高脂肪饮食(等热量、等氮营养成分),经过8天严格饮食再经过1~2周“洗脱”期后再进行自身交叉试验,与高碳水化合物饮食组相比,高脂饮食组哺乳期妇女的母乳唾液酸化HMOs显著降低,岩藻糖基HMOs无明显变化。
选择植物性膳食还是动物性膳食对HMOs是否有影响,在一项评估纯素食、部分素食和非素食膳食对哺乳期妇女的母乳HMOs组成的研究发现,这三类饮食的母乳样本中总HMOs成分没有显著性差异,且总的岩藻糖基HMOs和唾液酸化HMOs含量也无显著性差异[30]。哺乳期妇女饮食中的微量营养素含量对HMOs是否有影响,现阶段的证据也有限。一项研究中发现饮食中维生素A摄入量较高的哺乳期妇女的母乳中唾液酸化HMOs浓度较高[31]。
总的来说,评估母亲膳食摄入与HMOs含量和分布的相关研究较少,且评估方法的差异太大,膳食与HMOs之间的关系仍处于推测状态,怀孕期间的饮食摄入和食物的质量似乎对HMOs没有显著影响[27],而一些单独的饮食成分和补充剂可能短期对HMOs有影响[28]。
3.4 体质量指数与HMOs
在分析孕前体质量指数(body mass index,BMI)与HMOs是否相关时,研究结果之间出现了一些分歧。Ferreira等[32]发现孕前BMI与LNnT浓度呈正相关,但Samuel等[24]发现,孕前BMI与LNnT、LNT及LNFPV浓度呈负相关;同时在哺乳的前4个月,只有3′-SL和6′-半乳糖基乳糖(6′-Galactosyllactose,6′-GL)与孕前较高的BMI呈正相关。Larsson等[33]的研究结果却并未发现孕前BMI与这些HMOs之间的关联。可能因为这些研究是在不同的队列中进行的,遗传因素或地理位置存在比较大的影响。
在分析产后BMI与HMOs是否相关时,研究结果比较一致,总HMOs和总岩藻糖基化HMOs含量与产后BMI均呈正相关,特别是2′-FL。对于单个HMOs与产后BMI的关联,McGuire等人[34]发现产后BMI和与2′-FL和FLNH呈正相关,与DSLNT和LNnT呈负相关。Isganaitis等[35]发现,产后1个月的BMI与2′-FL、LNFP I显著负相关,与LNFPII和LNFP III显著正相关,但这些相关性并未延伸至产后6个月。Larsson等[33]发现,产后5个月的BMI与6′-SL和LSTb呈负相关,与2′-FL呈正相关。总之,需要更多的证据来证实母亲BMI与单一HMOs之间的关联。
3.5 其他因素
除了以上因素外,母乳中HMOs的含量可能还受到环境、胎龄、分娩方式等因素的影响。非洲冈比亚的一项人群研究报道,与在旱季哺乳时相比,在雨季哺乳的母亲产生的HMOs浓度较低[36]。此外,加拿大的一项研究表明,气候、阳光和过敏原暴露等其他季节性因素可能会影响加拿大哺乳期妇女的HMOs合成[27]。胎龄方面,与足月产后的母乳相比,早产后的母乳中含有更高的3′-SL、LSTb、DSLNT、LSTa、LSTc和更低的6′-SL[37-38]。剖宫产产妇的母乳中2′-FL、3′-SL、6′-GL浓度低于阴道分娩产妇的母乳,剖宫产和阴道分娩期间母体产生的生理差异,如应激反应、激素分泌等可能是HMOs组成产生差异的原因[38]。
4 总结与展望
HMOs的含量在母乳固体成分中仅次于乳糖和脂肪。越来越多的证据表明,HMOs对婴幼儿健康有着重要作用,HMOs有益生元、病原体抑制、抗炎、免疫调节和促进大脑发育等作用。母乳中HMOs的含量和种类受泌乳阶段、遗传因素、饮食、BMI、环境、胎龄和分娩方式等因素的影响。但是目前对于HMOs的研究方法缺少“金标准”,各研究中母乳采集的时间和方式,以及HMOs的检测和分析方法各有不同,且研究对象样本量相对较少,使得在获取标准化HMOs数据时出现一定困难,影响进一步的研究结果。因此,优化研究方法、扩大样本量将使我们能够更好地了解影响母乳中HMOs分布的各种因素,而后我们希望通过调节相关因素优化HMOs分布进而促进婴儿的健康水平。此外,HMOs对婴儿的临床意义及影响机制尚未完全明确,未来需要更多相关方面的研究进行深入探讨。
