粗糙脉孢菌发酵豆渣产物的品质和抗氧化活性
2023-03-06赵秀芳姚海燕袁江兰康旭
赵秀芳,姚海燕,袁江兰,康旭
(湖北工业大学生物工程与食品学院,湖北 武汉 430068)
豆渣是豆浆、豆腐等豆制品加工的副产物。我国每年可产生2 000多万吨湿豆渣[1],但新鲜豆渣水分含量高达80%[2],不仅口感粗糙,而且极易酸化腐败,因此,大部分豆渣经过直接干燥后用作动物饲料,也有一部分被直接丢弃[3]。分析表明,豆渣营养丰富,每100 g干豆渣含蛋白质约19.32 g、碳水化合物51.80 g、脂肪12.40 g,此外还含有大豆异黄酮、无机盐、单糖、低聚糖和B族维生素等[4]。豆渣中还含有多种抗营养因子,如凝集素和胰蛋白酶抑制剂等,直接食用可能引起腹泻或胃肠紊乱,利用微生物发酵处理可以有效去除这些抗营养因子,因此微生物发酵可能会大幅提升豆渣品质,丰富豆渣的用途。
近年来有关豆渣发酵的研究较多,例如利用马氏克鲁维酵母发酵豆渣,豆渣粗脂肪、可溶性膳食纤维和多糖分别增加24.5%、15.8%和26.2%,植酸和胰蛋白酶抑制剂活性分别降低61.7%和92.7%,豆腥味明显减少[5]。利用黑曲霉对豆渣进行发酵,氨基酸态氮含量为未发酵豆渣的3.23倍,纤维素酶活提升了2.02倍,半纤维素酶活提高了10.64倍[6]等。粗糙脉孢菌(Neurospora crassa)属真菌门、脉纹孢菌属,是粗壮串珠霉的有性世代,是一种能够产生纤维素酶的可食用真菌[7],有气生菌丝,能产生黄色或橙红色孢子。该菌能够分解豆渣中部分粗纤维和大分子蛋白质,产生小分子单糖、双糖、低聚糖、多肽、氨基酸等生理活性物质,使豆渣口感变细腻,提高豆渣的营养价值和综合加工特性。酿酒酵母具有较好的发酵特性,其利用谷物发酵基质产生酶系,对谷物的营养和功能品质产生较大影响[8];植物乳杆菌是工业生产中应用较多的发酵菌,具有优良益生保健功能和产细菌素的特性[9],但在豆渣发酵中应用较少。本研究以粗糙脉孢菌为基础发酵菌种,考察其与酿酒酵母、植物乳杆菌耦合对豆渣品质和抗氧化功能的影响,为豆渣类功能食品或食品、饲料、医药原辅料的开发提供参考。
1 材料与方法
1.1 菌种及主要试剂
新鲜豆渣:武汉远大豆制品有限公司;粗糙脉孢菌(Neurospora crassa):广州纳豆生物科技有限公司;酿酒酵母(Saccharomyces cerevisiae,菌种编号 CICC 1477):湖北安琪酵母有限公司;植物乳杆菌(Lactobacillus plantarum,菌种编号CICC 24809):北京川秀国际贸易有限公司;干酪素、羧甲基纤维素钠、1,1-二苯基-2-三硝基苯肼(1,1-diphenyl-2-trinitrophenylhydrazine,DPPH)、α-淀粉酶(100 000 U/g):上海源叶生物科技有限公司;福林酚、乙二胺四乙酸二钠(ethylenediamine tetraacetic acid disodium salt,EDTA-2Na)、邻苯三酚:国药集团化学试剂有限公司;3,5-二硝基水杨酸、5-磺基水杨酸:上海麦克林生化科技有限公司;考马斯亮蓝(G-250):西安飞扬生物科技有限公司;牛血清白蛋白(bovine serum albumin,BSA):德国BioFroxx公司;胰蛋白酶(猪胰)(250 U/mg):Aladdin试剂(上海)有限公司;18种氨基酸标准品:美国Sigma公司;其它试剂均为国产分析纯。
1.2 仪器与设备
DHWS-250型低温恒温恒湿培养箱:合肥达斯卡特生物科技有限公司;UPI-11-5T型优普系列超纯水器:四川优普超纯科技有限公司;Bio-Rad Mini-PRO TEAN Tetra Cell蛋白电泳装置:美国伯乐公司;K9860全自动凯氏定氮仪:山东海能科学仪器有限公司;L-8900型全自动氨基酸分析仪:日本Hitachi公司;UV-2600型紫外可见分光光度计:岛津仪器(苏州)有限公司;CR-13型柯尼卡美能达色谱仪:日本大阪市美能达有限公司;Agilent 7890A/5975C型气相色谱-质谱联用仪:美国Agilent公司。
1.3 试验方法
1.3.1 发酵用孢子粉制备
将粗糙脉孢菌接种至马铃薯葡萄糖琼脂培养基(potato dextrose agar,PDA)中,在 30℃恒温培养箱中培养48 h,收集孢子粉置于4℃冰箱备用。
1.3.2 发酵豆渣样品制备
粗糙脉孢菌发酵豆渣:称取含水量约为85%豆渣,100℃常压蒸20 min,冷却后置于超净台上,紫外灭菌15 min,按照豆渣质量0.01%接入粗糙脉孢菌孢子粉,搅拌均匀后置于30℃、相对湿度85%的恒温恒湿培养箱中发酵,记为NF。以未发酵豆渣作为对照。
接入0.01%粗糙脉孢菌和0.01%酿酒酵母粉,记为NSF,其余制备条件相同。
接入0.01%粗糙脉孢菌孢子粉和0.01%植物乳杆菌菌粉,记为NLF,其余制备条件相同。
接入0.01%粗糙脉孢菌孢子粉、0.01%酿酒酵母粉和0.01%植物乳杆菌菌粉,记为NSLF,其余制备条件相同。
1.3.3 豆渣固态发酵前后纤维素酶活的测定
参考文献[10]的方法测定纤维素酶活。分别取10 g新鲜发酵 0、24、36、48、60 h 的豆渣样品于烧杯或试剂瓶中,充分捣碎,再加入200 mL预冷的pH5.0乙酸-乙酸钠缓冲液(含0.15 mol/L NaCl),冰浴搅拌提取1 h,低温过滤,取1 mL滤液用相应的缓冲溶液适度稀释后测定纤维素酶活。纤维素酶活单位定义:在(40±1)℃、催化反应的pH值条件下,每分钟从5 mg/mL羧甲基纤维素钠溶液中释放1 μg还原糖所需要的酶量为一个酶活力单位,结果以U/g表示。计算公式如下。
式中:c为试样反应溶液中葡萄糖的上机浓度,mg/mL;c0为试样空白反应溶液中葡萄糖的上机浓度,mg/mL;n为试样溶液定容后的稀释倍数;m为试样质量,g;t为酶反应时间,min;100 为试样定容体积,mL;180.2为葡萄糖摩尔质量,g/mol;1 000为转化因子。
1.3.4 膳食纤维含量测定
参考GB 5009.88—2014《食品安全国家标准食品中膳食纤维的测定》和文献[11]中的方法测定发酵和未发酵豆渣不溶性膳食纤维(insoluble dietary fiber,IDF)和可溶性膳食纤维(soluble dietary fiber,SDF)含量。
1.3.5 豆渣理化指标测定
将未发酵豆渣(unfermented okara,UF)即发酵0 h样品和发酵 24、36、48、60 h 的 NF、NSF、NLF、NSLF 样品冻干磨粉并过60目筛。不同发酵时间还原糖的测定采用 3,5-二硝基水杨酸(3,5-dinitrosalicylic acid,DNS)法[12];发酵60 h样品水解氨基酸和游离氨基酸含量参考文献[13]中的方法测定。
1.3.6 聚丙烯酰氨凝胶电泳(polyacrylamide gelelectrophoresis,PAGE)
参考文献[14]中的方法,分别称取一定质量发酵60h的豆渣蛋白至10mL离心管,未发酵豆渣蛋白为对照,溶于0.01 mol/L pH7.4磷酸盐缓冲液中,保证蛋白浓度为10 mg/mL,将离心管在滚轴混匀仪上以50 r/min混样1 h,使蛋白充分溶解,稀释5倍,8 000 r/min离心15 min,取上清液备用。将上清液和含有巯基乙醇的上样缓冲液按照体积比1∶1混合,煮沸5 min,冷却后10 000 r/min离心2 min,取10 μL上样,分离胶和浓缩胶浓度分别为12%和4%。
1.3.7 总酸测定
参考GB 12456—2021《食品安全国家标准食品中总酸的测定》中的方法对新鲜发酵60 h豆渣样品进行总酸测定。
1.3.8 色度测定
不同发酵时间样品分别冻干磨粉并过60目筛,参考文献[15]中的方法测定 L*、a*、b*。
1.3.9 挥发性成分测定
采用顶空固相微萃取(headspace solid phase microextraction,HS-SPME)与气相色谱-质谱联用法(gas chromatography-mass spectrometry,GC-MS)[16]。取 7.5 g(精确至0.001 g)新鲜豆渣发酵60 h产物,置于40 mL顶空样品瓶中,在80℃条件下平衡30 min,使用50/30 μm 二乙烯基苯/碳分子筛/聚二甲基硅氧烷(divinylbenzene/carboxen/polydimethylsiloxane,DVB/CAR/PDMS)纤维萃取头进行固相微萃取,顶空吸附30 min,解吸5 min,每个样品重复测定3次。
GC条件:CP-SIL 8型毛细管柱(30 m×0.32 mm,0.25 μm),升温程序:40℃保持 2 min,先以 5℃/min升温到120℃保持2 min,再以7℃/min的速率升到220℃保持5 min;载气(He)流速1.0 mL/min,进样口温度250℃,不分流。
MS条件:采集方式为选择离子扫描,溶剂延迟7 min,质量范围35 amu~395 amu,离子源温度230℃,四极杆温度150℃,接口温度250℃,电离方式电子电离(electron ionization,EI),离子源能量 70 eV。
对匹配度大于70(最大值100)的挥发性成分根据NIST 08数据库对化合物进行定性,面积归一化法计算相对含量。
1.3.10 抗氧化活性测定
取不同发酵时间(36、48、60h)的豆渣干粉各2g,加入30 mL去离子水,28℃水浴搅拌1 h,然后8 000 r/min离心15 min获得上清液,用去离子水稀释20倍后待测。DPPH·清除率参考文献[17]中的方法并稍作修改;羟基自由基(·OH)清除率参考文献[18]中的方法测定;超氧阴离子自由基(·O2-)清除率参考文献[19]中的方法并稍作修改。
1.4 数据处理
采用Origin 2017作图,用SPSS 23.0对数据进行统计,采用单因素方差分析中的Duncan多重比较法进行显著性分析(p<0.05表示差异显著)。
2 结果与分析
2.1 豆渣发酵过程纤维素酶活变化规律
豆渣发酵过程中的纤维素酶活变化见图1。
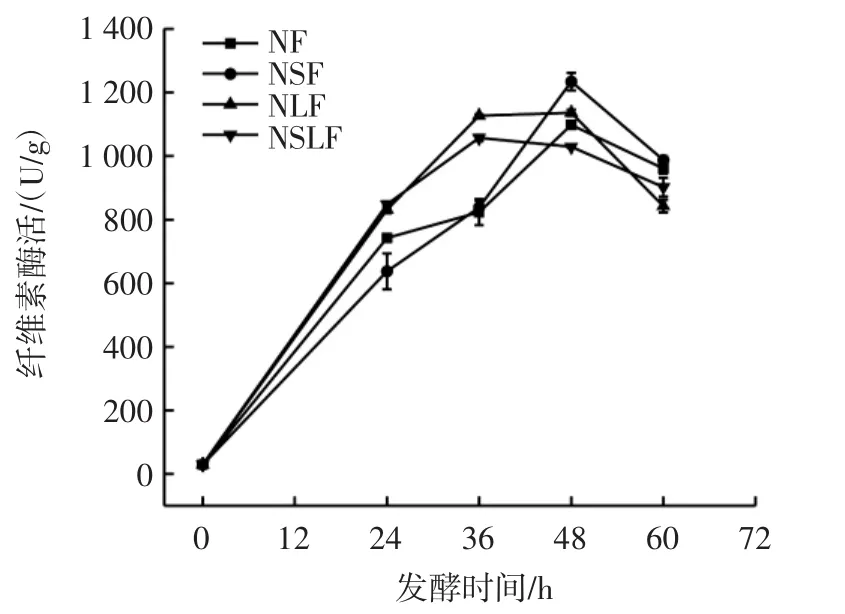
图1 豆渣发酵过程中的纤维素酶活Fig.1 Cellulase activity during okara fermentation
由图1可知,4组发酵豆渣样品的纤维素酶活均呈现先升高后降低的趋势,其中NF和NSF纤维素酶活均在发酵48 h时达到峰值,分别为1 097.98 U/g和1 234.18 U/g。而NLF和NSLF均在36 h达到峰值,分别为1 126.70 U/g和1 056.94 U/g,随后纤维素酶活逐渐降低。纤维素酶活能够影响发酵产物中膳食纤维的含量,纤维素酶可以将基质中不溶性膳食纤维分解成单糖,从而导致不溶性膳食纤维含量降低,可溶性膳食纤维含量增加。纤维素酶主要由粗糙脉孢菌产生[20],结果表明,酿酒酵母和植物乳杆菌参与发酵明显加快了粗糙脉孢菌的生长和产酶进程,发酵后期可能因为pH值明显降低,微生物的生长进入衰亡期,逐渐停止产酶,因此纤维素酶活下降。
2.2 发酵对豆渣膳食纤维和还原糖含量的影响
不同发酵豆渣中膳食纤维和还原糖含量差异见图2。

图2 不同发酵豆渣中膳食纤维和还原糖含量差异Fig.2 Differences of dietary fiber and reducing sugar contents in different fermented okara
由图2A可知,未发酵豆渣(UF)中IDF含量最高,占豆渣干重69.85%,SDF含量最低,为4.05%。菌种耦合发酵 60 h后,NF、NSF、NLF、NSLF 样品中 IDF 与未发酵豆渣相比均显著降低(p<0.05),分别为55.04%、43.27%、52.11%、41.09%,SDF含量明显提高。NSLF组IDF含量降低明显,推测酿酒酵母和植物乳杆菌能够协同促进粗糙脉孢菌产生更多纤维素酶,导致IDF被分解得更多,这与2.1中纤维素酶活变化规律基本一致。
由图2B可知,随着发酵时间延长,还原糖含量逐渐增加,而NSF组在发酵后期还原糖含量有明显下降趋势。在发酵36 h~48 h酿酒酵母参与的发酵有利于还原糖的积累,含量最高分别达到了(59.50±2.45)mg/g,继续发酵至终点(60 h)时,还原糖降低为53.40 mg/g左右。粗糙脉孢菌在发酵过程中产生的大量纤维素酶作用于发酵基质中的IDF,分解产生低分子的还原糖,因此还原糖含量增加,IDF含量降低,SDF含量增加。NLF和NSLF组还原糖积累量较低,这是由于植物乳杆菌消耗还原糖用于自身生长繁殖的速率高于酿酒酵母[21]。
2.3 不同发酵豆渣氨基酸差异分析
发酵豆渣水解氨基酸组成和含量变化见表1。发酵豆渣游离氨基酸组成和含量见表2。
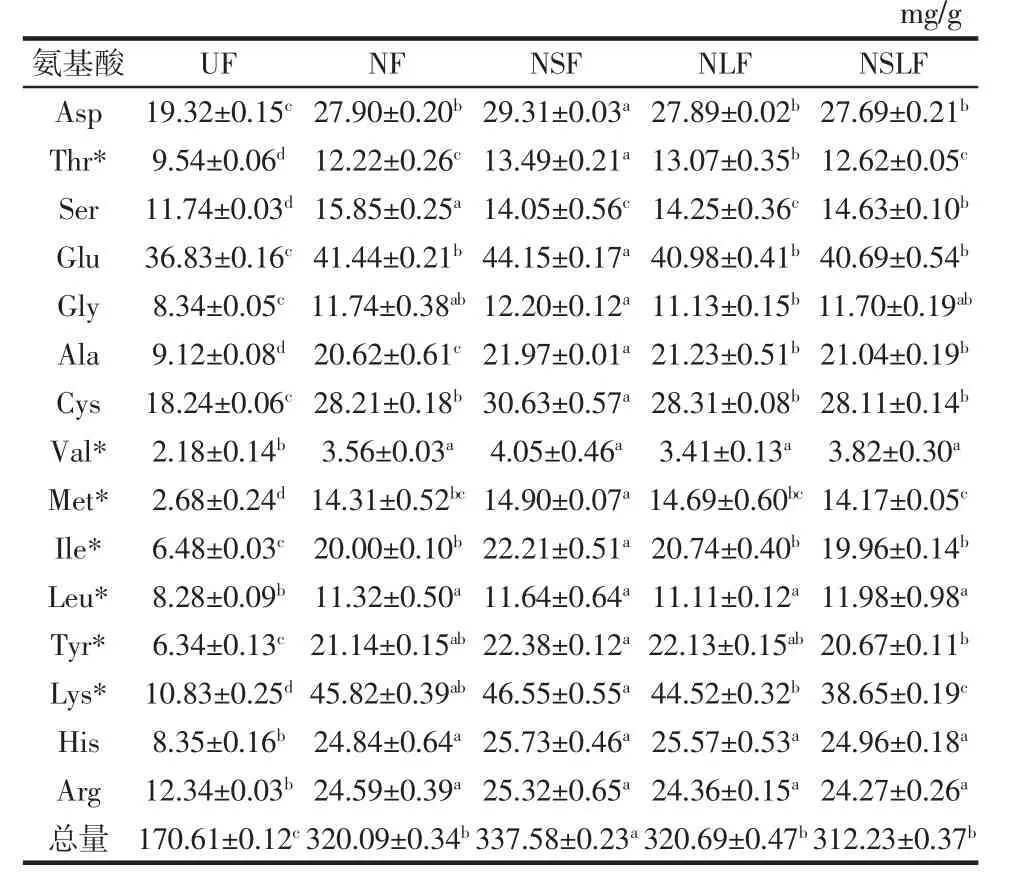
表1 发酵豆渣水解氨基酸组成和含量变化Table 1 Composition and content of hydrolyzed amino acids of fermented okara

表2 发酵豆渣游离氨基酸组成和含量Table 2 Composition and content of free amino acids of fermented okara
由表1、表2可知,发酵后4组豆渣的水解氨基酸以及游离氨基酸均显著增加(p<0.05)。NF、NSF、NLF、NSLF水解氨基酸含量分别为UF的1.87、1.99、1.88、1.83倍,必需氨基酸含量分别为128.37、135.22、129.67、121.87 mg/g,较 UF(46.33 mg/g)分别提高了 63.91%、65.74%、64.27%、61.98%,经过发酵后豆渣中必需氨基酸含量均能达到水解氨基酸的20%以上,豆渣营养品质明显提高。
由表2可知,UF游离氨基酸总量为(2.24±0.07)mg/g,而4组发酵豆渣的游离氨基酸明显增加,分别为 UF 的 13.16、10.93、16.97、14.69 倍,其中以NLF组增量最大。食品中呈鲜味氨基酸主要是谷氨酸和天冬氨酸,甜味氨基酸主要来源于丙氨酸和甘氨酸等。NLF组谷氨酸含量最高为2.07 mg/g,是UF的12.94倍;NLF组丙氨酸含量最高约为1.10 mg/g,是UF的6.88倍。这一结果表明植物乳杆菌参与的发酵对豆渣的滋味和营养提升作用更加明显。发酵促进蛋白质的降解,明显促进豆渣内呈结合状态活性物质的溶出与释放。水解氨基酸和游离氨基酸含量的变化与Vong等[22]的研究结果基本一致。
2.4 SDS-PAGE分析
发酵60h的豆渣蛋白SDS-PAGE电泳如图3所示。

图3 发酵豆渣蛋白的SDS-PAGE电泳Fig.3 SDS-PAGE of fermented okara protein
由图3 UF条带可以看出,豆渣中主要有7S和11S两种抗原蛋白[23]。7S蛋白包括α、α'、β三种亚基,分子量从上到下分别为82 kDa的α'亚基、71 kDa的α亚基、55 kDa的β亚基,11S蛋白包括38 kDa和30 kDa的酸性亚基、22 kDa的碱性亚基。与未发酵豆渣相比,4种发酵豆渣样品7S蛋白均明显发生降解,几乎看不到条带,11S蛋白38 kDa条带酸性亚基降解较充分,11S蛋白亚基的降解程度明显低于7S蛋白亚基。发酵后豆渣中抗原蛋白消失或含量减少,豆渣的致敏性明显降低。各发酵样品在10 kDa~15 kDa出现颜色较深的新条带,这可能是由于微生物将大分子蛋白质逐步降解为低分子蛋白质。与Kunitz型胰蛋白酶抑制剂(KTI)条带比较可知,发酵豆渣中胰蛋白酶抑制剂基本可以忽略。
2.5 发酵豆渣总酸含量
发酵豆渣总酸含量见图4。
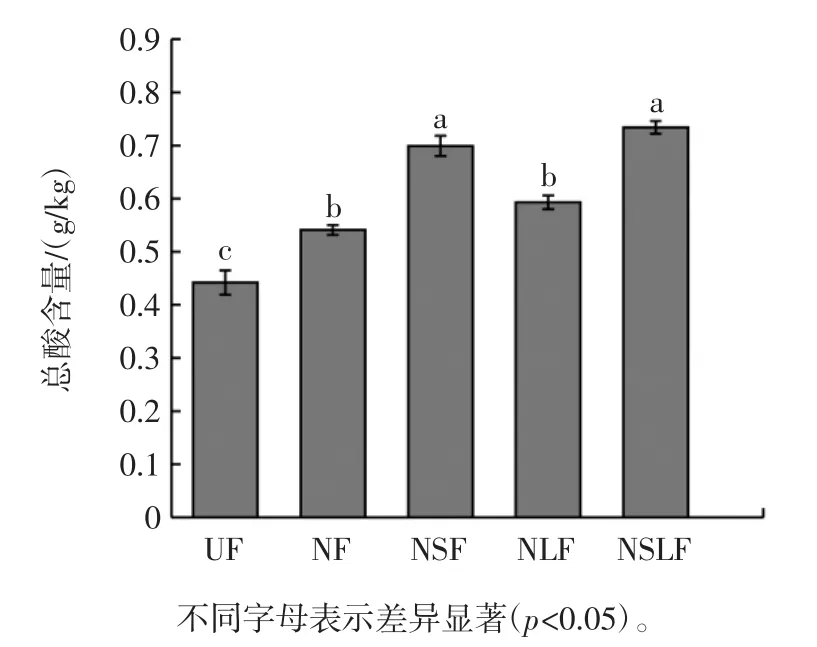
图4 发酵豆渣总酸含量Fig.4 Total acid content of fermented okara
由图4可知,发酵60 h后豆渣总酸含量相比未发酵豆渣均显著增加(p<0.05),分别可达 0.541、0.699、0.593、0.734 g/kg,酿酒酵母能利用豆渣基质中营养物质产生二氧化碳和乙醇,促进系统产生更多酶,因此有利于产生更多的有机酸、脂肪酸、氨基酸等酸性物质[24],使总酸含量显著增加(p<0.05)。植物乳杆菌经过发酵主要产生乙酸、乳酸等有机酸,比较NLF和NF总酸含量发现,植物乳杆菌的引入对总酸含量的增加无明显促进作用。
2.6 发酵豆渣感官质量差异分析
2.6.1 色度分析
豆渣发酵期间色度变化见表3~表5。

表3 豆渣发酵期间色度L*的变化Table 3 Change of chroma L*in okara during fermentation

表4 豆渣发酵期间色度a*的变化Table 4 Change of chroma a*in okara during fermentation
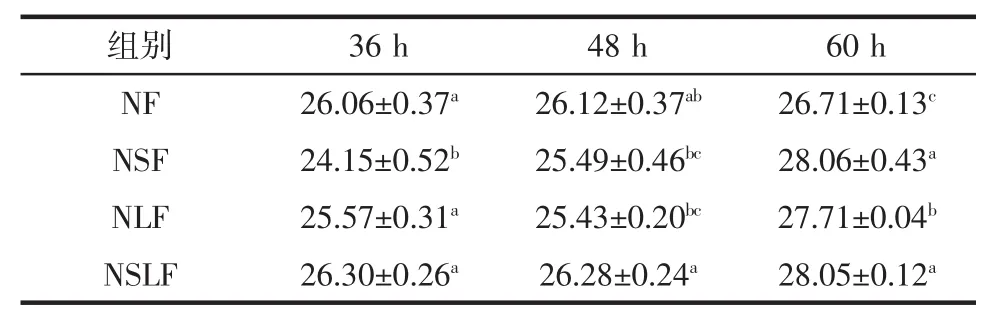
表5 豆渣发酵期间色度b*的变化Table 5 Change of chroma b*in okara during fermentation
由表3~表5可知,4组 L*均在68.27~74.26,均有较好的明亮度,这与粗糙脉孢菌的基本发酵特征有关。在本试验条件下,经过观察发现,粗糙脉孢菌在发酵前期产生肉眼可见的白色菌丝,发酵36 h时开始产生大量鲜亮的橙黄色孢子,从而导致发酵豆渣的L*均有明显增加,即明度增加。NF、NSF、NLF组L*随着发酵时间的延长而降低,NSLF组的L*在整个发酵过程均低于其它3组(除60 h)。
耦合发酵60 h的样品红色度(a*)明显高于粗糙脉孢菌单菌种发酵样品,特别是酿酒酵母参与耦合发酵尤为明显,以3个菌种耦合发酵的样品红色度增加最快,最终红色度最高。发酵36 h时NSLF组较另外3组呈现出显著差异(p<0.05),红色度最高。4组样品在发酵36 h时均能产生大量橙黄色孢子,并且红色度随着发酵时间的延长而增加,这与粗糙脉孢菌孢子富含类胡萝卜素有关。4组样品的黄色度(b*)表现出同红色度(a*)相似的变化趋势。
2.6.2 挥发性香气成分
发酵60 h豆渣挥发性香气成分分析见表6,不同种类挥发性成分相对质量分数见图5。
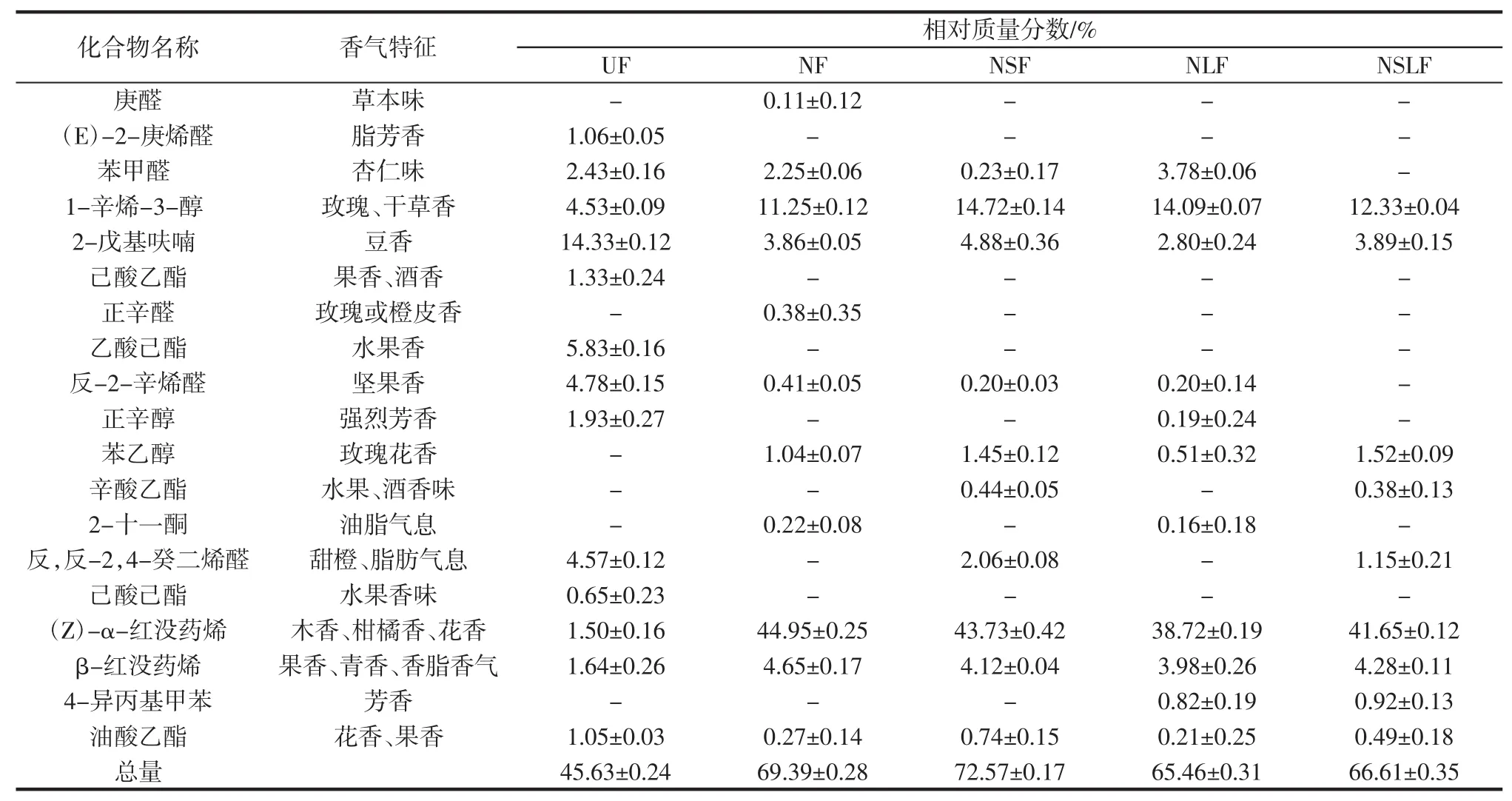
表6 发酵60 h豆渣挥发性香气成分分析Table 6 Analysis of volatile aroma components of okara fermented for 60 h

图5 不同种类挥发性成分相对质量分数Fig.5 Relative mass fractions of different volatile components
由表6可知,发酵后挥发性香气总量明显增加,NSF组最高为72.57%,是UF组的1.59倍。NF组(Z)-α-红没药烯含量最高为44.95%,是未发酵豆渣的30倍,该物质具有柑橘香、花香等风味。庚醛具有草本气息、正辛醛具有玫瑰或橙皮香气,这2种成分均仅在NF发酵组产生。发酵后1-辛烯-3-醇的含量明显提高,NSF样品最高,为14.72%,是UF的3倍,该物质具有令人愉悦的玫瑰和干草香气。苯乙醇在未发酵样品中未检出,经过发酵后NSLF组含量最高为1.52%,其具有清甜的玫瑰花香味,酵母细胞能够通过莽草酸途径形成的苯丙酮酸脱羧脱氢形成苯乙醇[25]。苯甲醛、苯乙醇、辛酸乙酯等成分含量的增加也对发酵后豆渣香气有贡献。辛酸乙酯具有水果香味,在有酿酒酵母的发酵组出现,该成分在NSF组含量最高,为0.44%,酯类成分是由于酿酒酵母的作用下醇与酸的酯化反应产生的,由于酯类较低的气味阈值[26],其在较低含量时也能被感知。
由图5可知,发酵豆渣中挥发性成分种类主要有醛类、醇类、酯类、烷烃类、芳香烃等。经过发酵后,醇类、烯烃类和芳香烃类挥发性成分相对含量明显增加,醇类最高为NSF组21.96%,是UF的4倍,烯烃类最高为NF组50.52%,是UF的14倍。醛类、酯类、烷烃类和杂环类挥发性成分相对含量有所降低,推测酯类含量的减少可能是因为菌种利用酯类等营养物质进行自身代谢将其转化成醇类物质等。豆渣在菌种作用下经过长时间发酵发生一系列生化反应,如羟醛缩合、蛋白质降解、醇酸缩合等[27],产生出多种具有特殊香味的挥发性物质,这些化合物互相融合,使发酵产物具有独特的风味。因此,粗糙脉孢菌协同酿酒酵母和植物乳杆菌耦合发酵对豆渣风味具有明显提高作用。
2.7 菌种耦合发酵对豆渣抗氧化活性的影响
豆渣发酵期间抗氧化活性的变化见图6。

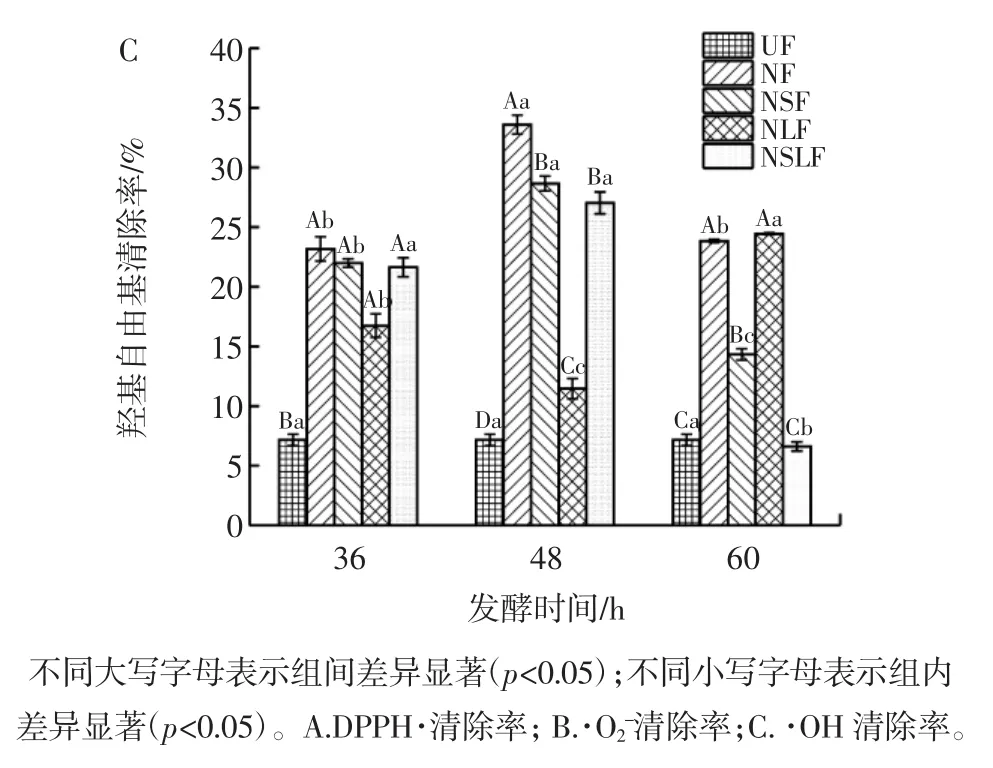
图6 豆渣发酵期间抗氧化活性的变化Fig.6 Changes in antioxidant activity during soybean dregs fermentation
由图6可知,发酵后豆渣DPPH·清除率均能达到95%左右,而未发酵豆渣DPPH·清除率测定值仅为13%左右,表明发酵前豆渣没有明显的DPPH·清除能力,而各发酵样品均具有很强的DPPH·清除能力,发酵后豆渣中还原糖含量明显增加,还原糖可能提供电子使自由基形成稳定的物质,有效阻断自由基链式反应[28],豆渣抗氧化能力明显提高。·O2-清除率也比未发酵豆渣有显著提高(p<0.05)。NF、NSF、NLF 三组的·O2-清除率均较高,并在发酵36 h达到峰值,分别为74.44%、86.03%及83.79%,之后随着发酵时间延长而降低。从·OH清除率来看,在发酵 48 h时,NF、NSF、NSLF组·OH清除率达到最高值,发酵后期·OH清除率下降,可能是由于还原类物质在后期被微生物用于自身生长消耗。发酵豆渣过程中可能产生了类胡萝卜素和抗氧化活性多肽等物质[29],从而提高了豆渣的抗氧化能力,但需要进一步试验确认。
3 结论
本文以粗糙脉孢菌为基本发酵菌种,耦合酿酒酵母和植物乳杆菌发酵鲜豆渣,探究豆渣的营养品质和抗氧化活性的变化,以获得营养丰富、感官品质良好的功能性食品、饲料或医药辅料。豆渣发酵后,还原糖、总氨基酸、游离氨基酸、可溶性膳食纤维等功能活性物质含量明显增加,豆渣的营养组成更有利于人体健康。NSLF组IDF被微生物明显分解,IDF含量降低28.76%,SDF含量提高5.03%。植物乳杆菌参与的发酵对豆渣的滋味提升效果最好,粗糙脉孢菌协同酿酒酵母和植物乳杆菌耦合发酵能够有效增加挥发性成分的种类并提高含量,发酵产物风味更加优良。发酵后豆渣的DPPH·清除能力、·O2-清除能力和·OH清除能力均明显增加,抗氧化活性明显增强。综合分析认为,粗糙脉孢菌、酿酒酵母和植物乳杆菌耦合发酵豆渣能够明显改善豆渣的营养品质和抗氧化活性,本研究中豆渣经过发酵后营养成分明显改善、豆渣利用价值有效提高,为开发以发酵豆渣为原辅料的功能性食品提供参考。
