果香型葡萄酒酵母的发酵特征与转录组学研究
2023-02-17许维娜李记明刘延琳
许维娜,王 磊,李记明*,刘延琳
(1 西北农林科技大学 陕西杨凌 712100 2 烟台张裕集团有限公司 山东省葡萄酒微生物发酵技术重点实验室 山东烟台 264001 3 滨州医学院药学院(葡萄酒学院) 山东烟台 264003)
酵母菌是葡萄酒生产中的主要微生物,对葡萄酒的风味及质量至关重要[1]。 葡萄酒发酵过程中,酵母负责将葡萄汁中的糖转化成酒精和CO2,并生成数百种香气物质,形成葡萄酒最重要的香气——发酵香(酒香、二类香气),与品种香(果香、一类香气)和陈酿香(三类香气)一起构成葡萄酒复杂而独特的风味[2-3],因此,酵母被称为“葡萄酒之父”。
葡萄酒中的主要香气物质有酯类(乙酸酯类和脂肪酸乙酯类)、醇类、酸类、酚类、酮类、醛类、萜烯类、含硫化合物等[4]。 其中部分在葡萄原料中是以结合态存在的潜在香气物质,需要酵母代谢进行转化和激发。 目前葡萄酒中发现的千余种香气物质,超过400 种是在酵母发酵过程中产生的[5]。性状优良的果香型酵母能充分激发葡萄原料中的潜在香气,对提升葡萄酒果香型风味与质量是必不可少的[6]。我国气候以及葡萄种植多样化的特点造就了丰富的葡萄酒酵母资源[7]。由于长期的自然选择作用,酵母菌株已经逐渐适应了我国的风土和葡萄特性,这些本土酵母用于葡萄酒酿造时,能够充分反映葡萄品种的特色,具有强化葡萄酒中风土特性的潜力,酿造出个性和风格突出的优质葡萄酒[8-9]。 本土优良酵母的选育及应用对于国产葡萄酒质量提升具有重要意义。
本项目组前期从陕西蛇龙珠发酵样品中分离到一株果香表现突出的酿酒酵母菌株SSF12 (专利号ZL 2020 1 1494317.7),将其与另一株果香表现较差的菌株各自发酵后,收集酵母细胞进行转录组测序,研究两株酵母菌转录组学的差异。结合发酵液的理化指标、香气物质含量和感官特性,解析果香型酵母代谢机理,为优良酵母菌开发提供参考依据。
1 材料与方法
1.1 材料与试剂
野生葡萄酒酵母SSF12、NGF6 为前期从陕西蛇龙珠、 宁夏贵人香发酵的葡萄酒样品中分离所得;蛇龙珠葡萄取自烟台蓬莱某葡萄园。
葡萄糖、蛋白胨、酵母粉、异丙醇、无水乙醇、氯仿,国药集团化学试剂有限公司;Takara 酵母RNAiso plus 试剂盒,Takara 公司;DNase-free 水、RNase-free 水,金克隆(北京)生物技术有限公司。YEPD 培养基:葡萄糖20 g/L,蛋白胨20 g/L,酵母粉10 g/L。
1.2 仪器与设备
M391F 垂直层流超净工作台,法国Erlab 公司; 发酵罐,烟台帝伯仕自酿机有限公司;5180R台式大容量离心机,德国Eppendorf 公司;UVS-99核酸蛋白测定仪,上海彦哲仪器设备有限公司;Agligent 6890N-5975B 气相色谱-质谱联用仪,安捷伦科技有限公司。
1.3 试验方法
1.3.1 酵母的活化、接种、发酵 配制YEPD 培养基,进行SSF12 和NGF6 的划线转接与活化。
成熟的蛇龙珠葡萄除梗破碎,浸渍12 h 后将皮渣过滤除去,取纯汁。用葡萄汁进行酵母种子液的制备,以5%的接种量接种至20 L 发酵罐中,24~26 ℃恒温发酵。每日监测发酵液比重和失重情况。
1.3.2 葡萄酒基础理化指标的测定 待发酵液比重≤0.995 且日失重趋于稳定时,停止发酵(发酵6 d)。将发酵液澄清、倒罐并于4 ℃下储藏。对酒样的基本理化指标进行检测,包括还原糖、总糖、总酸以及酒精度。 还原糖、 总糖测定采用费林滴定法,总酸测定采用酸碱滴定法,酒精度测定采用密度瓶法[10]。 每个酒样设3 个重复。
1.3.3 香气成分与含量的检测 将2 g 氯化钠和8 mL 酒样放入萃取瓶中并密封。 添加混合内标用于半定量分析(内标1:薄荷醇464.5 μg/L;内标2:2-辛醇500 μg/L)。 将样品在45 ℃平衡5 min 后,用SPME Arrow 自动进样装置以250 r/min 的转速下平衡样品,并在相同转速下用SPME Arrow 纤维进行50 min 吸附。 待吸附平衡后迅速将进样器插入气相色谱进样口解析,检测色谱柱:WAX 毛细管柱(60 m×0.25 nm×0.25 nm);程序升温:初始温度40 ℃,保持1 min,以3 ℃/min 升温至180℃,再以20 ℃/min 升温至230 ℃,保持15 min;载气(He):1.2 mL/min;进样口温度:250 ℃,压力:20.0 kPa。 质谱条件:传输线温度280 ℃,离子源温度230 ℃,溶剂延迟0.5 min。 每个样品设2 个平行。将各香气成分质谱图结果与NIST2008 标准谱库进行比较,结合人工图谱解析,进行香气物质定性;采用2-辛醇内标法进行定量[11-12]。
1.3.4 感官质量分析 感官分析小组由15 名经过专业训练的品酒师组成(7 男8 女,22~51 岁),参照GB/T 15038-2006[10]中的感官分析方法,从果香香气(40 分)、果香口感(40 分)、果香整体(风格和典型性,20 分) 三个方面对不同酒样进行感官质量评价。 每个酒样设3 个重复。
1.3.5 发酵样品中酵母的收集与转录组学测序发酵6 d 后,离心收集酵母细胞 (6 000 r/min,4℃,5 min),并用DNase-free 水洗涤2 遍。 每5×107个细胞中加入1 mL 的RNAiso Reagent 溶液,根据试剂盒说明进行RNA 的提取。 获得RNA 沉淀后,加入RNase free 水使其完全溶解。用紫外吸收法检测RNA 的浓度与纯度(A260/280≥1.8)。 质检合格的RNA 样品送至上海美吉生物医药科技有限公司进行Illumina 转录组学测序。 每个样本设2个平行。
获得原始数据后,先进行过滤,提高其质量。过滤结束后,以酿酒酵母S.cerevisiae S288C(GCF_000146045.2_R64)的基因组为参考,采用TopHat2 (http://ccb.jhu.edu/software/tophat/index.shtmL)进行比对分析。采用RSeQC(v2.6.3)对测序饱和度、基因覆盖率和重复读取进行分析,以评估转录组数据的总体质量。 为了鉴定差异基因(Differentially expressed genes,DEGs),采用FPKM(Fragments per kilobase of exon per million mapped reads)计算每个基因的差异表达水平,并使用EdgeR (Empirical analysis of Digital Gene Expression in R,v3.10)进行差异表达分析,将Padjust<0.05 且|log2fold change (FC)|≥1 设置为默认阈值。 根据Ding 等[13]的方法,将DEGs 用Kyoto Encyclopedia of Genes and Genomes(KEGG)进行功能注释与富集分析。
2 结果与分析
2.1 葡萄酒样品基本理化指标
两酵母发酵样品的基本理化指标如表1 所示。 SSF12 发酵的样品酒精度相对较高,糖含量和总酸较低,而NGF6 发酵的样品酒精度较低,糖含量和总酸较高。 总体而言,SSF12 发酵样品的发酵程度和糖利用率更高。

表1 发酵样品的基本理化指标Table 1 Basic physicochemical parameters of fermentation samples
2.2 香气物质测定结果
两样品中的香气物质检测结果如表2 所示,共检测出73 种香气物质,按照结构特性将其分为醛酮类(6 种)、酯类(44 种)、醇类(9 种)、酸类(8种)、萜烯、C13-降异戊二烯类(5 种)、酚类(1 种)等。
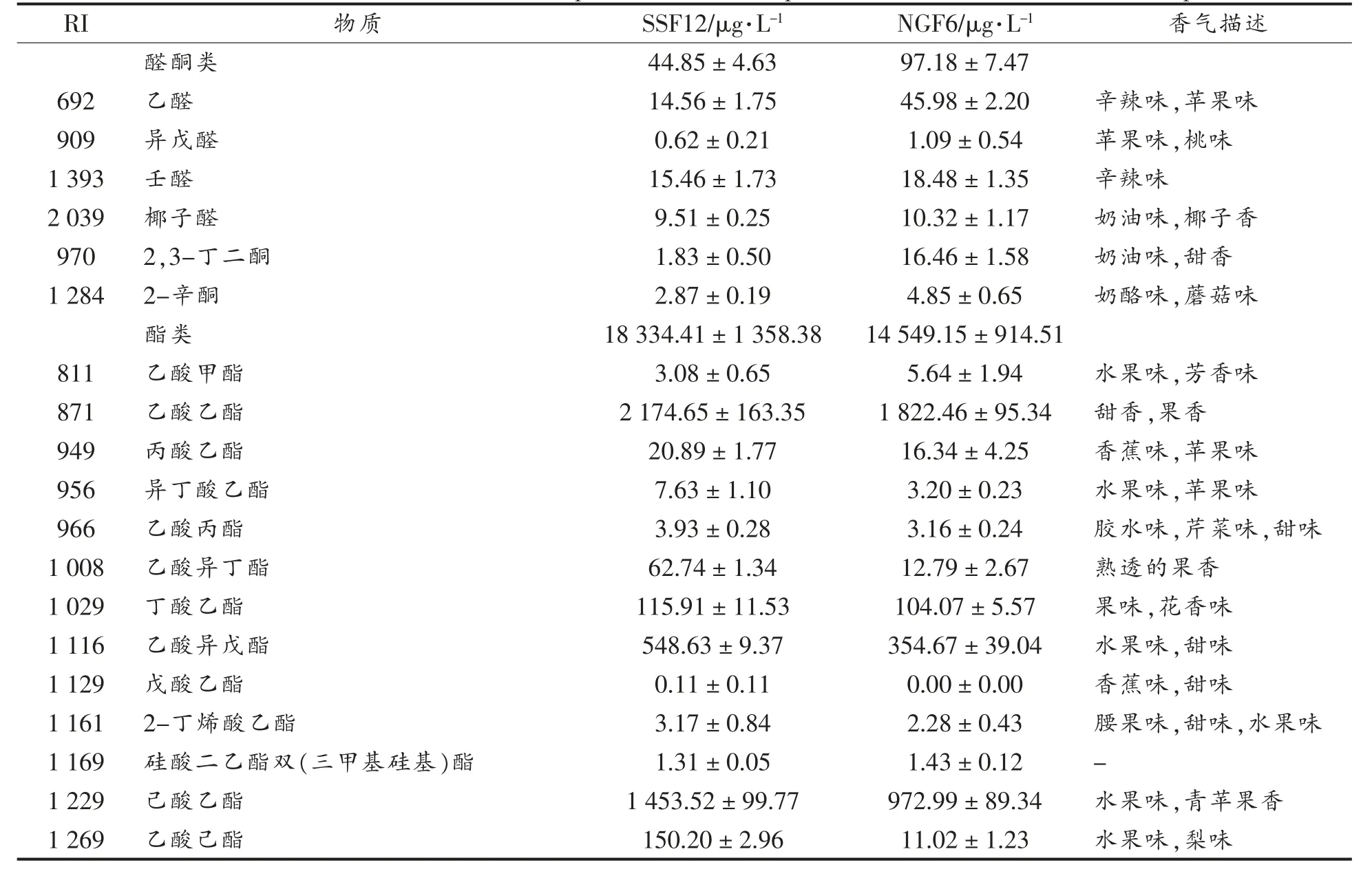
表2 发酵样品香气成分含量与香气描述Table 2 The content and aroma description of aroma components in different fermentation samples
两样品中,酯类物质的种类和含量均最多,醇类、 酸类物质次之,酚类最少。 戊酸乙酯只在SSF12 的发酵样品中检测出来,其它物质在两样品中均存在。
检测到的香气物质中,种类和含量最多的为酯类,大多为葡萄酒贡献果香、花香[14]。SSF12 发酵样品中酯类物质含量较NGF6 更高,预期花果香更明显。含量第二的是醇类,通常来源于葡萄浆果本身以及发酵过程[15]。 低浓度的高级醇(<300 mg/L)能够增加葡萄酒香气的复杂度[16],两样品中醇类物质含量均较为适宜,SSF12 略高。 葡萄酒中不同浓度的脂肪酸类、 醛类物质会给酒体带来不同的影响,适宜浓度时有助于塑造酒体的层次感和丰富度,浓度过高时则会产生腐败和刺激性气味[17-18]。 两样品中酸类物质含量均在适宜范围内,且差别不大(NGF6 中含量略高);醛类物质虽浓度有差异(NGF6 中含量更高),但也均在适宜范围内。 另外,SSF12 中萜烯、C13-降异戊二烯类、酚类物质含量也略高于NGF6。 总体而言,SSF12 的香气物质种类和含量均高于NGF6。
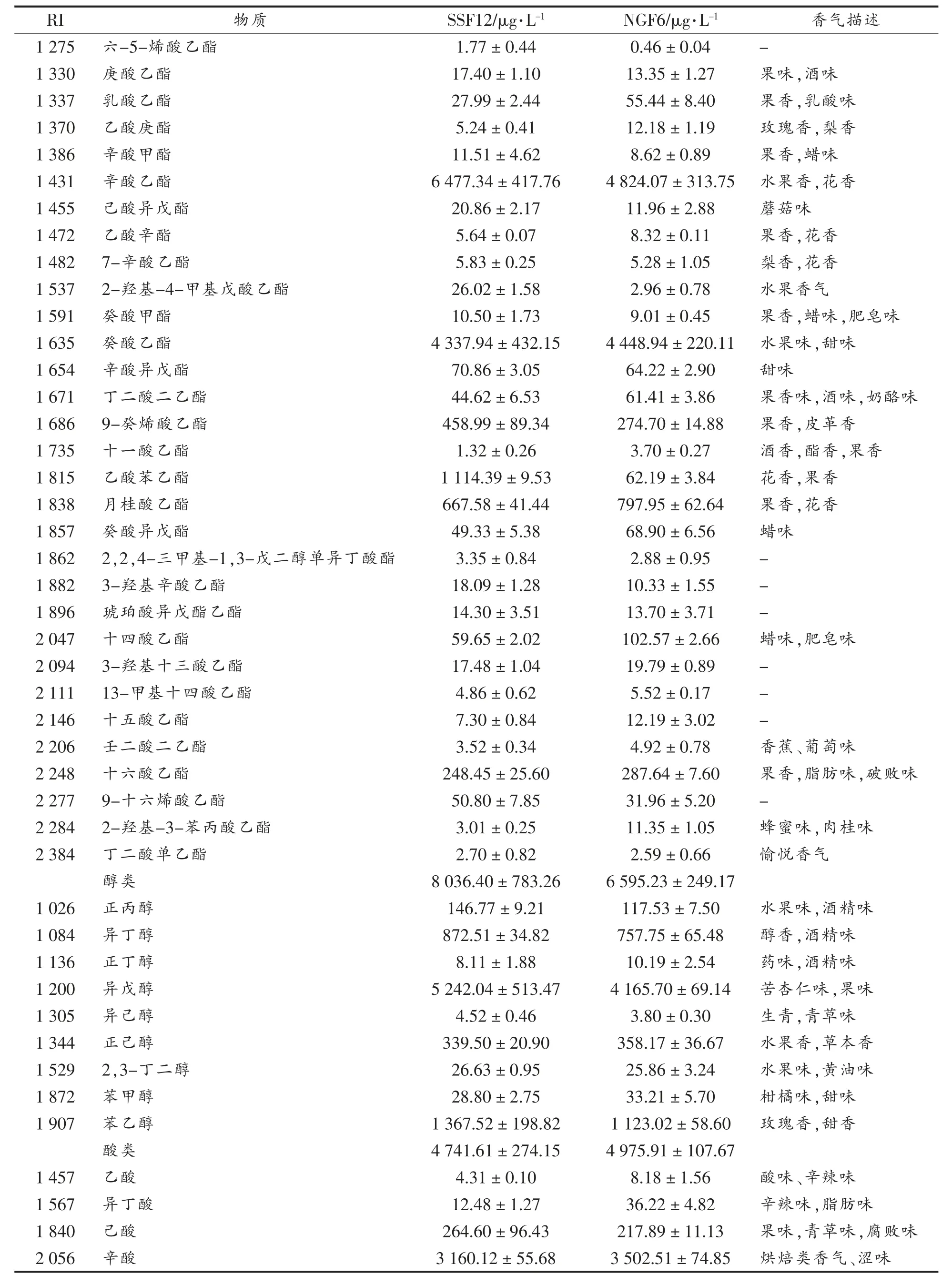
(续表2)

(续表2)
2.3 感官质量分析结果
两样品的感官分析结果如表3 所示,其外观无明显差异,SSF12 发酵的样品口感柔和,果香香气和口感更为突出,酒体更加平衡、协调,综合质量更佳。

表3 感官质量品评数据Table 3 Aroma sensory evaluation data
2.4 酵母转录组学分析结果
利用Illumina HiSeq 技术对两酵母进行转录组学分析,筛选|log2FC(SSF12/NGF6)|≥1 且Padjust<0.05 的基因作为差异基因(DEGs),共检测出了434 个差异基因,包括290 个上调基因 (图1,红色)和144 个下调基因(图1,绿色)。
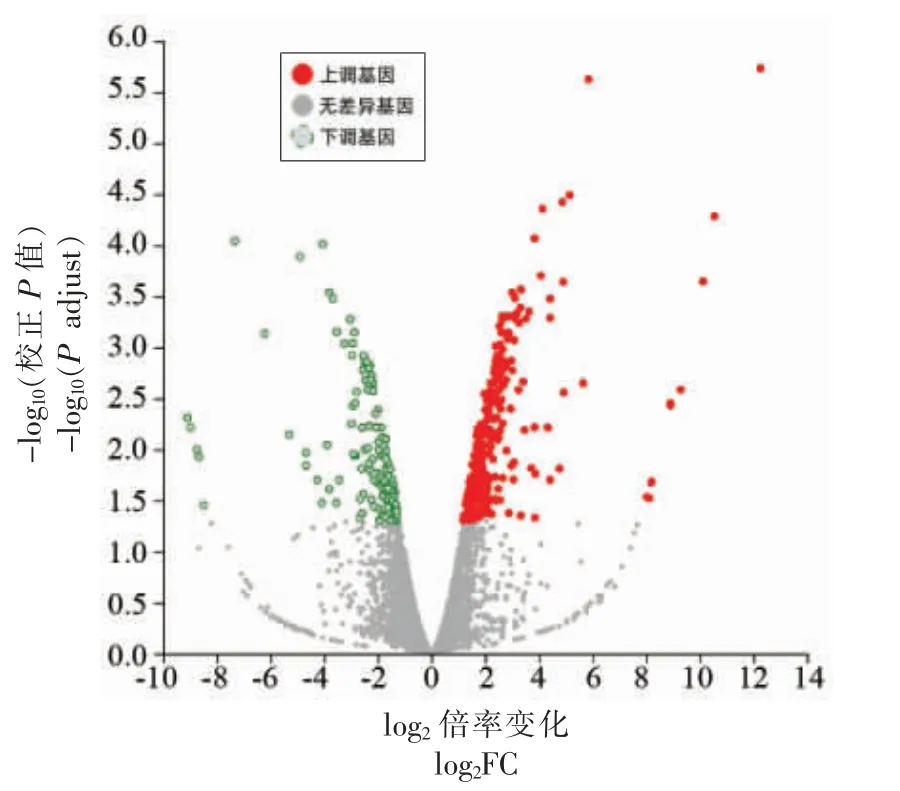
图1 SSF12 与NGF6 的基因表达差异火山图Fig.1 The volcano graph of gene expression difference between SSF12 and NGF6
2.4.1 差异基因的KEGG 聚类分析结果 采用KEGG 聚类分析对果香酵母SSF12 和非果香型酵母NGF6 之间主要代谢途径的差异进行表征,结果如图2 所示。 所有的DEGs 被划分为5 个主要类别,包括代谢(碳水化合物代谢,核苷酸代谢,氨基酸代谢,辅因子和维生素代谢,能量代谢,脂质代谢6 个亚类,141 个DEGs),遗传信息处理(转录,翻译,折叠、 分类和降解3 个亚类,69 个DEGs),细胞过程(细胞生长与死亡,运输和分解代谢2 个亚类,34 个DEGs),环境信息处理(信号转导1 个亚类,9 个DEGs)和生物体系统(环境适应1 个亚类,4 个DEGs)。 大部分途径中上调基因比下调基因的数量更多,只有氨基酸代谢与能量代谢途径中包含更多的下调基因,表明SSF12 中碳水化合物(包括葡萄糖、氨基糖和核苷酸糖、乙醛酸和二羧酸、丙酮酸、磷酸戊糖、淀粉和蔗糖、半乳糖、果糖、甘露糖等)、辅因子(泛酸、CoA、叶酸、核黄素)、核苷酸等的代谢效率更高,并且SSF12在DNA 转录、翻译、细胞生长、物质运输、信号转导、环境适应等方面表现更好,这些现象可能有助于SSF12 快速适应发酵环境[19];而NGF6 在部分氨基酸(如酪氨酸、苯丙氨酸、组氨酸、精氨酸等)和氮代谢(能量代谢)方面有一定优势。

图2 差异基因的KEGG 聚类统计柱形图Fig.2 Statistics histogram of KEGG pathways of the DEGs
2.4.2 中心碳代谢及香气物质代谢途径分析 葡萄酒发酵过程中的香气物质产生情况是酵母菌的重要特征之一,这些香气物质主要包括脂肪酸、高级醇、酯类、醛类、氨基酸等[20-21],它们大多是中心碳代谢途径的次级产物。 为了探明果香型酵母中心碳代谢和香气物质代谢的情况,对这些途径及相关DEGs 进行了更详细的研究,结果如图3 所示。
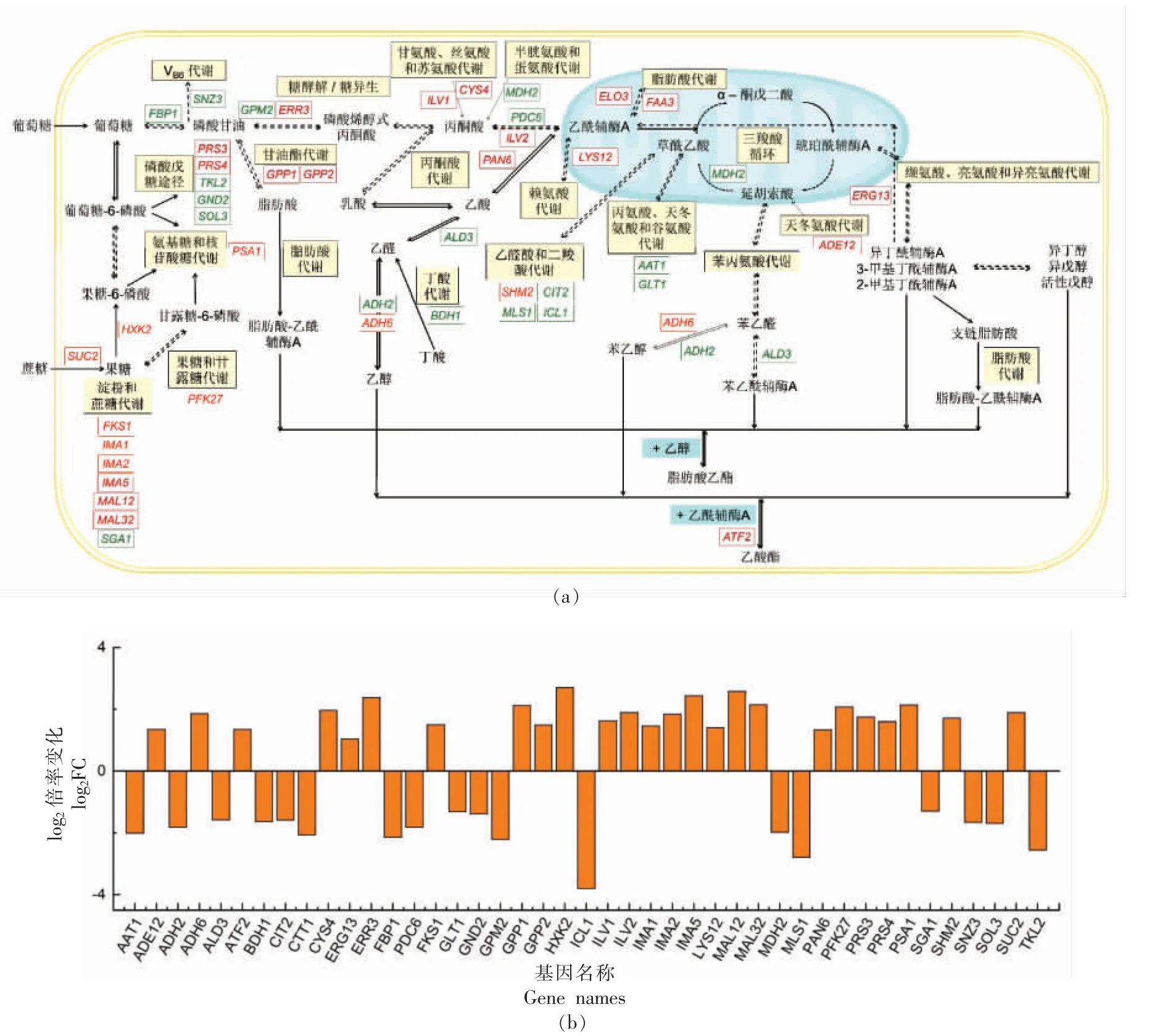
图3 中心碳代谢和香气物质代谢途径中的差异表达基因(a)及差异表达倍数(b)Fig.3 The transcriptome profiles (a) and fold changes (b) of DEGs involved in the central carbon metabolism and volatile compound metabolism
淀粉和蔗糖代谢 (Starch and sucrose metabolism) 途径中,SSF12 有8 个上调基因和1个下调基因。 其中,只有编码葡聚糖1,4-α-葡萄糖苷酶的SGA1 基因表达下调,log2FC 值为-1.30,其它基因均上调,包括IMA1,IMA2,IMA5,MAL12,MAL32 等葡萄糖苷酶基因。 这些基因的集中转录上调有助于蔗糖、葡萄糖、果糖等糖类的相互转化。
糖酵解(Gluconeogenesis)途径是葡萄糖经过一系列的酶促反应生成丙酮酸的过程,乙醛、乙酸和乙醇均在糖酵解中产生。此代谢途径中,相较于NGF6,SSF12 有HXK2,ERR3,ADH6转录上调,FBP1,GPM2,PDC6,ADH2,ALD3 转录下调。 据报道,HXK2 参与葡萄糖抑制的调节,而且此调节作用具有很强的菌株特异性[22-23]。 乙醇脱氢酶(ADH家族)催化醛及相应醇的转化[24],其中乙醇脱氢酶Ⅵ(ADH6p)参与醇类的合成,Mendes 等[25]认为酿酒酵母发酵过程中ADH6 等基因的高水平表达与异丁醇、异戊醇和苯乙醇产量的提高相关,本研究中也观察到了类似现象 (表2)。 乙醇脱氢酶II(ADH2p)催化醇向醛的转化,刘奎等[26]研究发现中断ADH2 基因能够减少乙醇转化为乙醛,提高乙醇产率。 本研究中ADH2 的log2FC 值为-1.82,且SSF12 发酵液乙醇含量高于NGF6(表1),与文献报道结果相符。Pigeau 等[27]、Heit 等[28]报道ALD3编码的乙醛脱氢酶Ⅲ的高表达是导致冰酒中乙酸含量升高、乙酸乙酯含量降低的重要原因。本研究中NGF6 的ALD3 基因转录水平更高,乙酸含量高,乙酸乙酯含量低,这与文献报道相符。
磷酸戊糖途径 (Pentose phosphate pathway)是葡萄糖氧化分解的另一个途径,产生大量核糖和NADPH。 此途径中,SSF12 有PRS3 和PRS4 转录上调,TKL2、GND2、SOL3 转录下调。 PRS3 和PRS4 是编码磷酸核糖焦磷酸合成酶的5 个基因家族中的2 个,催化多种生物合成途径的第1 步,包括嘌呤和嘧啶的生物合成。 营养缺乏状态下,PRS3 对于维持细胞完整性、离子稳态和细胞骨架是必不可少的[29]。 同时,PRS3 的高表达有助于细胞提高对高乙酸等环境逆境的适应能力[30]。 GND2编码磷酸戊糖途径的关键酶6-磷酸葡萄糖酸脱氢酶,SOL3 编码的6-磷酸葡萄糖醛酸酶也是磷酸戊糖途径的重要酶类。 Liu 等[31]报道GND2、SOL3 等基因的增强表达对于NADPH 的再生具有重要意义,以提供乙醛转化、脂肪酸合成等所需的辅助因子。 本研究中NGF6 发酵液中乙醛含量高于SSF12 (表2),GND2、SOL3 的转录上调并未加剧这两种物质的消耗。 而NGF6 中脂肪酸类物质含量高于SSF12(表2),提示该菌株中过多的碳通量转向脂质的合成,Silverman 等[32]的研究中也发现了类似现象。
三羧酸循环 (TCA cycle) 途径中,SSF12 有MDH2 转录下调,二羧酸代谢(Glyoxylate and dicarboxylate metabolism)途径中,SHM2 转录上调,ICL1、MLS1 转录下调,其中MDH2、ICL1、MLS1 的下调可能会减弱对乙醛酸循环和糖异生的回补[33]。另外,果糖和甘露糖代谢(Fructose and mannose metabolism)途径中,PFK27 转录上调;氨基糖和核苷酸糖代谢 (Amino sugar and nucleotide sugar metabolism) 途径中,PSA1 转录上调; 丁酸代谢(Butanoate metabolism) 途径中,ILV2 转录上调,BDH1 转录下调。研究报道,PFK27 编码6-磷酸果糖-2-激酶,是糖酵解途径的关键调节因子,会在葡萄糖缺乏的情况下抑制细胞生长[34];PSA1 的过表达能增强细胞对非适宜温度的耐受性[35];这两个基因的转录上调可能有助于增强SSF12 细胞的环境适应性。 ILV2 表达水平与高级醇产量负相关,即敲除该基因有助于提高高级醇含量[36],这与本研究SSF12 发酵液中高级醇含量更高的结果并不相符,这可能是由于基因转录与表达水平不一致,以及其它基因(ADH2、ADH6 等)转录调控的综合作用结果。
脂肪酸代谢中,参与甘油酯代谢(Glycerolipid metabolism) 的GPP1、GPP2 以及参与脂肪酸代谢(Fatty acid degradation/biosynthesis/elongation)的FAA3、ELO3 转录上调。 GPP1、GPP2 编码甘油-3-磷酸磷酸酶,催化甘油-3-磷酸向甘油的转化。 文献报道,GPP1 和GPP2 的高表达虽然不能显著增加甘油产量,但可增强细胞对渗透、厌氧和氧化应激的耐受性[37]。 FAA3 编码长链脂肪酸CoA 连接酶,ELO3 编码脂肪酸延长酶,这2 个基因的表达上调有助于脂质的积累。
另有多个DEGs 参与氨基酸代谢过程,如参与甘氨酸、丝氨酸和苏氨酸代谢的ILV1 和CYS4,参与赖氨酸合成的LYS12,参与丙氨酸、天冬氨酸和谷氨酸代谢的ADE12,参与缬氨酸、亮氨酸和异亮氨酸代谢的ERG13,以及参与β-丙氨酸代谢的PAN6 转录上调;参与丙氨酸、天冬氨酸和谷氨酸代谢的AAT1、GLT1,参与半胱氨酸和蛋氨酸代谢的MDH2 转录下调。 这些基因的转录调节可能影响相应氨基酸的合成和分解。
葡萄酒中由发酵产生的酯主要有两类,分别是乙酸酯(高级醇/乙醇和活性乙酰辅酶A 之间的缩合)和脂肪酸乙酯【活性脂肪酸(酰基辅酶A)/高级醇(酰基辅酶A)和乙醇之间的缩合】[38]。 ATF2(编码醇乙酰基转移酶Ⅱ,log2FC 为1.25) 参与乙酸酯的合成,其上调可能导致乙酸酯的合成代谢加快。本研究SSF12 发酵液中乙酸乙酯、乙酸异丁酯、乙酸异戊酯、乙酸己酯、乙酸苯乙酯等乙酸酯含量均高于NGF6(表2),可能与ATF2 的转录上调相关。
总的来说,SSF12 和NGF6 的中心碳代谢和挥发性化合物代谢途径存在差异。 SSF12 在糖类转化、高级醇生成、酯类代谢等途径中有更多的上调基因,而NGF6 在乙醛合成、脂肪酸合成等途径更具优势。
3 结论
对比果香型酵母SSF12 与果香表现较差的酵母NGF6 的发酵和转录组学数据,发现两菌株的代谢及代谢调控情况有较大的差异。 SSF12 糖利用度和发酵度高,发酵液中酯类、醇类、萜烯类物质含量高于NGF6,醛类、脂肪酸含量低于NGF6,果香感官特性明显优于NGF6。 转录组学分析发现,SSF12 在碳水化合物(尤其是糖类、高级醇、酯类)代谢、核酸代谢途径中表现更突出,NGF6 在乙醛合成、脂肪酸合成、部分氨基酸代谢途径中表现更好。 ADH2,ADH6,ALD3,ATF2 等基因在SSF12中转录上调,且与香气物质代谢密切相关,可作为基因工程改良酵母的研究基因,为果香型酵母的选育提供参考。
选育性状优良的本土特色酵母是国内外的研究热点。 本文通过对比分析果香型表现好的酵母与果香型表现差的酵母的发酵表现与转录组学差异,了解果香酵母香气物质代谢通路调节情况,解析各基因在发酵过程中的作用及影响,为果香型酵母的筛选与改良提供依据,有助于高效筛选高品质本土酵母,酿造出更具风土特性的国产葡萄酒。
