高通量SNP芯片在牛体外早期胚胎染色体质量鉴定中的初步应用
2022-11-29胡智辉巩建飞王雅春赵善江朱化彬
胡智辉,王 欢,衡 诺,巩建飞,王 忆,王雅春,赵善江*,朱化彬*
(1.中国农业科学院北京畜牧兽医研究所,农业农村部动物遗传育种与繁殖(家禽)重点实验室,北京 100193;2.中国农业大学,北京100193)
牛胚胎生产与移植技术可以充分利用优秀种公牛和优秀母牛的遗传资源[1-2],获得大量具有高遗传品质的牛体内、外胚胎,对于良种母牛繁殖潜力的挖掘、快速扩繁以及种质资源保护具有重要的价值。经过不断的研究和完善,牛体外胚胎生产技术取得了巨大进步,并在生产中得到广泛应用[3-4]。国际胚胎移植协会最新数据显示,2020年全球共生产可移植牛体内胚胎约36万枚,与2019年相比总体下降。然而生产的牛体外胚胎总数却逐年攀升,2020年已超115万枚[5]。随着活体采卵-体外受精技术在我国乃至世界范围的兴起[6-7],体外胚胎生产正在成为牧场优质种质资源扩繁的主要技术手段。但是,目前体外胚胎技术产业化应用还存在成本高、移植妊娠率低等问题[8-9]。在过去的15~20年里,体外胚胎移植妊娠率低这一现状非但没有得到改善甚至还在继续下降[10-11]。
胚胎质量是决定胚胎移植后妊娠率高低的关键[12-13],如果能在植入前对胚胎进行质量鉴定,在排除一些自身存在染色体问题的胚胎基础上实现对胚胎生产性能的评估,那么将大大提高胚胎移植后的受胎率和犊牛的价值。研究发现,体外胚胎染色体异常导致的胚胎发育能力异常是影响体外胚胎妊娠率最根本的原因[5, 14]。染色体异常与胚胎质量、胚胎移植后妊娠率以及胎儿发育直接相关,同时也是导致早期胚胎死亡以及先天性缺陷疾病的主要原因[15]。当下,人类胚胎植入前检测技术已经发展到了较高的水平,可以实现无创检测,筛选优质胚胎进行移植以保证受胎率和后代质量[16-17]。但是,牛胚胎遗传价值或生产性能的测定还处于起步阶段,相应的微创胚胎取样技术、单细胞全基因组扩增和芯片检测等相关工作尚未见在国内开展。因此,本研究以牛体内外胚胎为研究材料,探究利用胚胎微创取样技术对胚胎进行生产性能测定的可行性,以期为进口遗传物质监管和胚胎质量鉴定提供一套可行的操作方案。
1 材料与方法
1.1 试验材料
本研究所用体内胚胎为实验室自有胚胎,体外胚胎为实验室利用离体卵巢通过体外受精所生产的胚胎,离体卵巢来自河北省廊坊市大厂屠宰场,精液来自山东奥克斯公司。
1.2 主要试剂与仪器
所有体外胚胎生产相关试剂均购自Sigma公司;MALBAC全基因组扩增试剂盒购自亿康医疗;胚胎培养皿购自Thermo公司。CO2培养箱购自Thermo公司;体式显微镜购自Nikon公司;胚胎切割仪购自AB公司。
1.3 体外胚胎生产
1.3.1 卵母细胞采集及体外成熟 将屠宰场卵巢置于28~30 ℃灭菌生理盐水中保存,2 h内送达实验室。用加有双抗的灭菌生理盐水清洗3遍。用配有18号无菌针头的真空蠕动泵抽取卵巢上3~6 mm 卵泡,在体视显微镜下用口吸管挑选胞质均匀、卵丘致密且包裹至少 3 层以上卵丘细胞的卵丘-卵母细胞复合体(COCs),置于 38.5 ℃,5% CO2,饱和湿度的 CO2培养箱中成熟培养 22~24 h。
1.3.2 体外受精 卵母细胞处理:将体外成熟 22~24 h 后的COCs用受精液洗涤3次后,移入受精液微滴中,准备进行体外受精。精液处理:将冷冻精液在 38 ℃ 左右的水浴锅中解冻,用洗精液洗涤精液,然后1 800 r·min-1离心 8 min,弃上清,重复两次;用洗精液重新悬浮精子沉淀;用血细胞计数器计算精子密度,调整精子密度至每毫升 2×106个。体外受精:将精液重悬液加入含有COCs的受精液中,置于 38.5 ℃、5% CO2、饱和湿度的培养箱中精卵共孵育 8~18 h。
1.3.3 体外胚胎培养 将受精后的受精卵脱去颗粒细胞后移入前期液中继续培养,48 h 后观察并记录受精卵的卵裂情况。之后移入后期液中继续培养,此后每 2 d 进行一次半量换液,体外培养 7~8 d 后,观察并记录囊胚的发育情况。
1.4 胚胎切割取样
用不含钙、镁的DPBS在平皿中做3个洗滴,将胚胎在洗滴中洗涤两次后移入最后一个洗滴,此时胚胎可牢固的贴在底部,将平皿置于胚胎切割仪的显微镜下,通过操作臂快速切取少量滋养层细胞。用口吸管将滋养层细胞吸出并移入提前配制好的胚胎裂解液中(注意吸取过程应尽可能少带液),每切割一枚胚胎要重新换洗液,同时用酒精擦拭切割刀片,裂解液可直接进行全基因组扩增或暂存于-80 ℃。切割后剩余的胚胎移入后期液中继续培养24 h,观察胚胎是否继续发育并计算切割发育率。
体外2细胞和8细胞切割取样方法及注意事项与胚胎取样基本一致,将胚胎透明带切开,推出卵裂球,吸取卵裂球放入胚胎裂解液中。
1.5 血液采集
选取11头母牛,使用含EDTA的采血管进行采血,全部采集牛尾根部血液,暂存于-20 ℃。
1.6 DNA提取及全基因组扩增
对于胚胎样品,使用亿康医疗的MALBAC全基因组扩增试剂盒。在 5 μL胚胎裂解产物中加入 30 μL 预扩增混合液,放入PCR 仪中进行预扩增。预扩增结束后在 35 μL 的预扩增产物中加入 30 μL 扩增混合液,放入PCR仪中进行指数式扩增。扩增结束后使用NanoDrop 核酸测定仪检测扩增后DNA浓度及纯度,将符合要求的DNA存于-80 ℃备用,并计算扩增成功率,即扩增后的DNA量大于1 000 ng且OD值在1.7~2.1之间。
对于血液样品,使用磁珠法进行全血基因组DNA提取,简单地说就是向全血中加入裂解液、异丙醇、磁珠悬浮液,将离心管放于磁力架上待磁珠完全吸附于离心管侧壁后彻底弃去溶液,使用漂洗液进行两次漂洗,使用洗脱液将DNA洗脱下来,存于-20 ℃备用。
1.7 芯片检测
进行DNA扩增、酶促片段化、沉淀和重悬。随后样本过夜孵育的过程中与BeadChip杂交,DNA经退火得到位点特异性的50-mer探针,与某种Infinium微珠类型共价偶联。Infinium XT工作流程继续酶基延伸赋予等位基因特异性,然后进行荧光染色。iScan系统检测微珠的荧光强度,llumina软件自动执行分析和基因型识别。
1.8 芯片数据分析
各组样品命名如下:采集的牛全血样品组命名为血液组(blood)、切取的半个体内胚胎组命名为1/2体内胚组(half-vivo),体内胚胎切取的滋养层组命名为体内滋养层组(vivo-tec)、体外胚胎切取的滋养层组命名为体外滋养层组(vitro-tec)、体外获得的2细胞切取的单个卵裂球组命名为体外2细胞组(2-cell)、体外获得的8细胞切取的单个卵裂球组命名为体外8细胞组(8-cell)。
首先通过R语言将各组所有SNP缺失位点筛选出来,将至少3个连续SNPs缺失认为染色体片段缺失[18]。为进一步筛选各组在全基因组扩增前染色体缺失情况,同时减小全基因扩增随机性导致的假片段缺失,试验最终以连续7个SNPs(含7个)缺失认为染色体片段缺失。
1.9 基因筛选、GO注释和KEGG功能富集分析
本研究使用ensembl数据库(asia.ensembl.org)中的biomart 模块将体内滋养层组和体外滋养层组中连续7个及以上SNPs缺失的片段中包含的基因筛选出来,将筛选出的基因使用R语言进行GO注释和KEGG功能富集分析。
1.10 芯片数据填充及生产性能预测
本研究使用Beagle 5.1将Illumina 100K Bovine Bead chip芯片数据填充至Illumina 150K Bovine Bead chip。整合两种不同密度芯片:100K芯片位点共计95 256个,150K芯片位点共计123 268个,根据位点名称进行匹配,重合位点共计64 958个。使用Plink (https: //www.cog-genomics.org/plink2)处理软件将原始文件转化为VCF (variant call format)格式。使用Beagle 5.1软件进行基因定向和填充过程,计算填充准确性。使用一步法进行基因组育种值预测,结果共有3项:产奶量、乳脂量以及乳蛋白量。
1.11 统计分析
试验数据采用“平均值±标准差”表示,各组数据应用SAS 9.2进行单因素方差分析及多重比较,不同字母代表组间差异显著。
2 结 果
2.1 胚胎切割及发育率
试验分别对体内囊胚、体外囊胚、体外2细胞和8细胞胚胎进行切割取样,分别切取体内胚胎1/2(half-vivo)、一小部分滋养层(vivo-tec),体外胚胎一小部分滋养层(vitro-tec)、体外2和8细胞胚胎的单个卵裂球(2-cell和8-cell组),切割结果如图1所示。本研究中,2-cell和8-cell组仅作为单细胞扩增组,用于验证优化扩增程序,其余各组切割取样后均继续培养,观察发育率。切割后发育率结果如图2所示,vivo-tec组和vitro-tec组的切割发育率均大于90%((94.4±5.6)%vs.(90.5±6.6)%),half-vivo组切割后的发育率为(22.2±14.7)%(2/9)。这说明切取半个胚胎对胚胎的发育有较大影响,而切取少量滋养层细胞对胚胎继续发育影响较小,符合胚胎微创取样要求,可用于后续试验。

A.体内囊胚;B.切割后的体内囊胚;C.体内囊胚切割后培养24 h;D.体外囊胚;E.切割后的体外囊胚;F.体外囊胚切割后培养24 h

图2 各组胚胎切割后体外培养发育率
2.2 全基因组扩增成功率
胚胎切割后各组样品通过MALBAC法进行全基因组扩增,当扩增后的DNA量大于1 000 ng且OD值在1.7~2.1之间时认为扩增成功。扩增成功率如图3所示,1/2体内胚组和体外2细胞组DNA扩增成功率为100%,与上述两组相比,体内滋养层组、体外滋养层组和体外8细胞组扩增成功率呈依次下降趋势,其中体外8细胞组扩增成功率仅(60.0±24.5)%,为各组最低。这说明在相同的扩增条件下,全基因组扩增成功率可能与起始DNA含量正相关,切割获取的起始DNA量越多,扩增成功率越高。

图3 不同组扩增成功率的差异
2.3 全基因组扩增后DNA量
全基因组扩增后各组DNA量如表1所示,1/2体内胚组DNA量最高(3 277.69±105.41),体外8细胞组最低(1 030.00±427.04)。与1/2体内胚组相比,体内滋养层组和体外滋养层组全基因组扩增后DNA量均显著下降 (P<0.05),且体外滋养层组全基因组扩增后DNA量显著低于体内滋养层组 (P<0.05),与体外2细胞组相比,体外8细胞组全基因组扩增后DNA量显著下降 (P<0.05)。

表1 各组全基因组扩增后DNA量
2.4 芯片检出率
如图4所示,与扩增后DNA量一致,各组芯片检出率呈现相同的规律,1/2体内胚组的检出率最高,可用于后续生产性能评估,其余各组依次下降。相较于1/2体内胚组,体内滋养层组和体外滋养层组的检出率均显著下降(P<0.05),且检出率均低于90%,芯片检出率无法满足后续生产性能评估要求,需要进一步进行芯片填充才能预测胚胎价值。体外2细胞组和8细胞组的检出率也均低于90%,且8细胞组检出率显著低于其余各组(P<0.05)。进一步对各组扩增后DNA量与芯片检出率进行相关分析发现,两者相关度为0.709 5(图5),存在较高的相关性。综上,起始DNA含量是影响DNA扩增和芯片检出率的关键,起始DNA含量越高(切割取样量越大),样品全基因组扩增效率和芯片检出率越高。
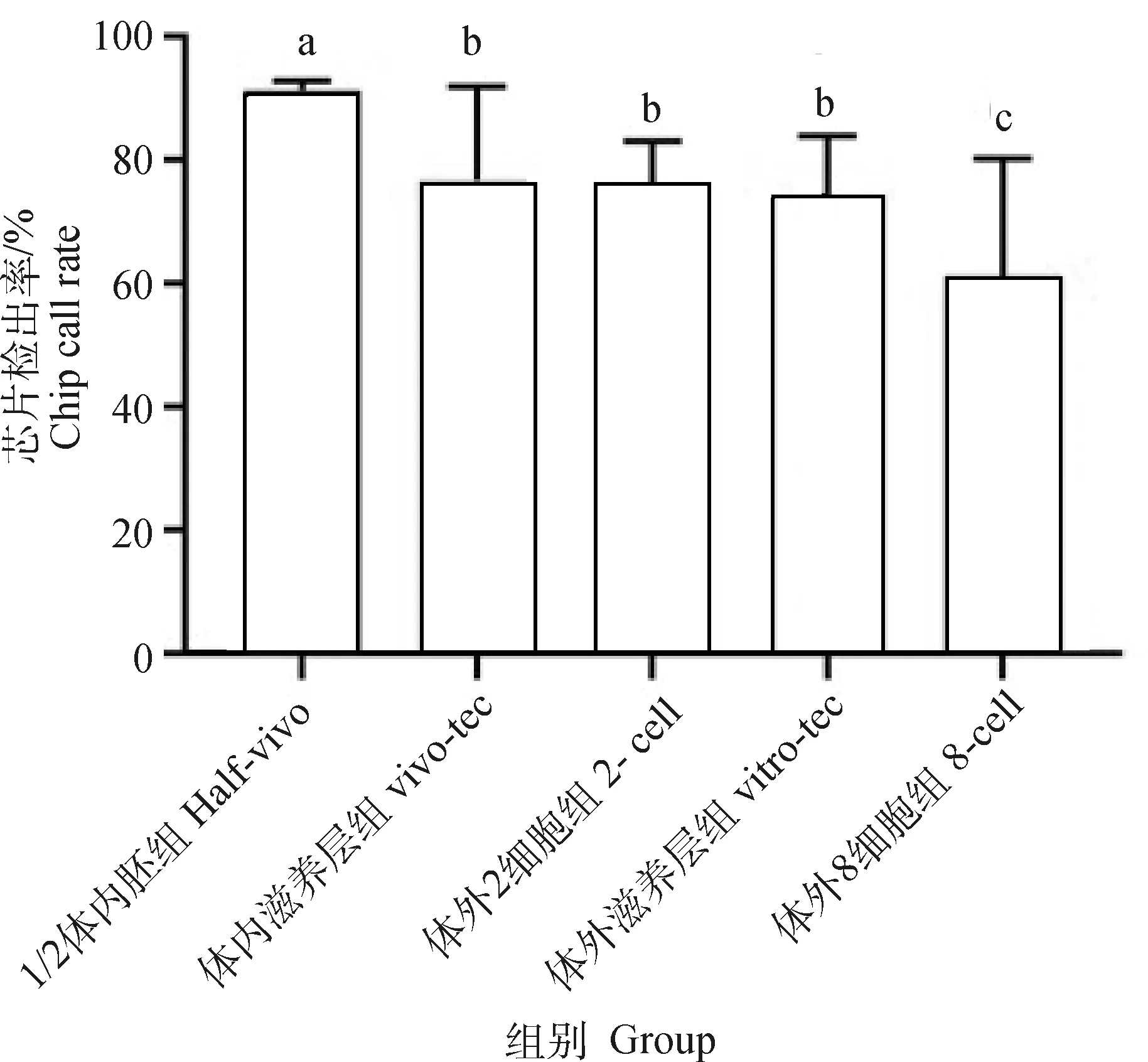
图4 不同组芯片检出率的差异
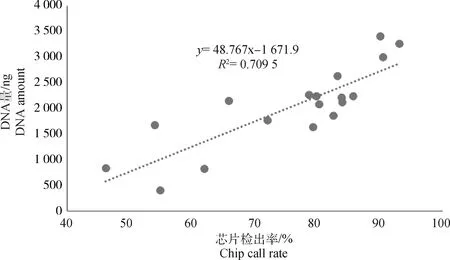
图5 DNA量与芯片检出率的关系图
2.5 不同组染色体片段缺失情况
为分析各组SNP缺失情况,尤其是检出率低于90%的各组缺失情况,进一步对各组30对染色体片段缺失情况进行量化统计。2细胞和8细胞在实际生产中不会用于胚胎取样,故本节重点分析体内、外滋养层组的染色体片段缺失情况。此外,为更准确地分析体内、外滋养层组染色体片段缺失情况,试验引入荷斯坦奶牛尾根血芯片数据(检出率均大于90%)作为试验对照组。对比分析1/2体内胚组、体内、外滋养层组与血液组在染色体片段缺失上的差异。
片段缺失标准参考Turner 等[18]的方法,以至少3个连续SNPs缺失认为染色体片段缺失,统计各组染色体片段缺失情况。结果如表2所示,血液组染色体片段缺失量最低(34),体内滋养层组最高(5 982),与血液组相比,1/2体内胚组染色体片段缺失量增加了8.9倍,而与1/2体内胚组相比,体内滋养层组染色体片段缺失量增加了16.9倍,体外滋养层组染色体片段缺失量增加了14.5倍。这说明滋养层取样会带来较大量的染色体片段缺失,而这一缺失是否会影响后续生产性能测定需要进一步加大试验研究量。

表2 不同组染色体片段缺失情况
为进一步筛选各组在全基因组扩增前染色体缺失情况,同时减小全基因扩增随机性导致的假片段缺失,试验进一步对染色体缺失片段所包含的连续SNPs数量进行了扩大统计,结果如表3所示,当连续缺失SNP≥7个时,1/2体内胚组不再存在片段缺失,故将染色体片段缺失所包含的SNPs数量扩大至7个用于后续试验分析。

表3 各组连续缺失7个及以上SNPs的片段数量
2.6 基因筛选、GO注释和KEGG功能富集分析
进一步分析了体外和体内滋养层组连续缺失≥7个SNPs的染色体片段,分别在体外和体内滋养层组发现了188和388个缺失片段。随后用ensembl数据库(asia.ensembl.org)中的biomart对筛选到的缺失片段进行筛选,结果显示在体外滋养层组发现46个基因,体内滋养层组发现48个基因。为了确定这些基因是否影响胚胎发育,对筛选到的基因进行GO和KEGG功能富集分析。GO富集分析结果显示,在体外和体内滋养层组分别获得340个和306个GO条目,其中排名前20的如图6所示(按P值排序)。通过KEGG富集分析,在体外滋养层组发现了17条显著的通路,在体内滋养层组发现了13条显著富集的通路。图7显示了TOP20的通路。

A.体内滋养层组;B.体外滋养层组

A.体内滋养层组;B.体外滋养层组
2.7 芯片填充准确性及育种值估计
本研究填充准确性是按等位基因一致性比率计算的,也就是正确填充基因型所占百分比,由R2表示。填充准确性分布可见图8,可以看出64 958个SNP填充位点中52 334个SNP填充位点的R2在0.99~1之间。本次填充的准确性为91%。填充后育种值见表4,表中可以看出体内滋养层组产奶量估计育种值最大值为203.94,最小值为61.69,平均值为127.68;乳脂量估计育种值最大值为3.36,最小值为0.88,平均值为2.39;乳蛋白量估计育种值最大值为1.13,最小值为-2.68,平均值为-0.84。体外滋养层组产奶量估计育种值最大值为55.59,最小值为-55.58,平均值为-6.07;乳脂量估计育种值最大值为9.04,最小值为-4.61,平均值为2.14;乳蛋白量估计育种值最大值为5.40,最小值为-3.75,平均值为0.47。从平均产奶量估计育种值看体内滋养层组远高于体外滋养层组,可能是因为体内胚胎在生产时选择的是产奶量较高的母本,而体外胚胎所选择的屠宰场卵巢并不知道生产性能,而且生产性能差的更容易被屠宰,这也说明了为什么体外滋养层组的产奶量普遍偏低且有正有负。而1/2体内胚组的母本为试验用牛且生产性能不高,所以获得的3枚胚胎产奶量、乳脂量、乳蛋白量都偏低。
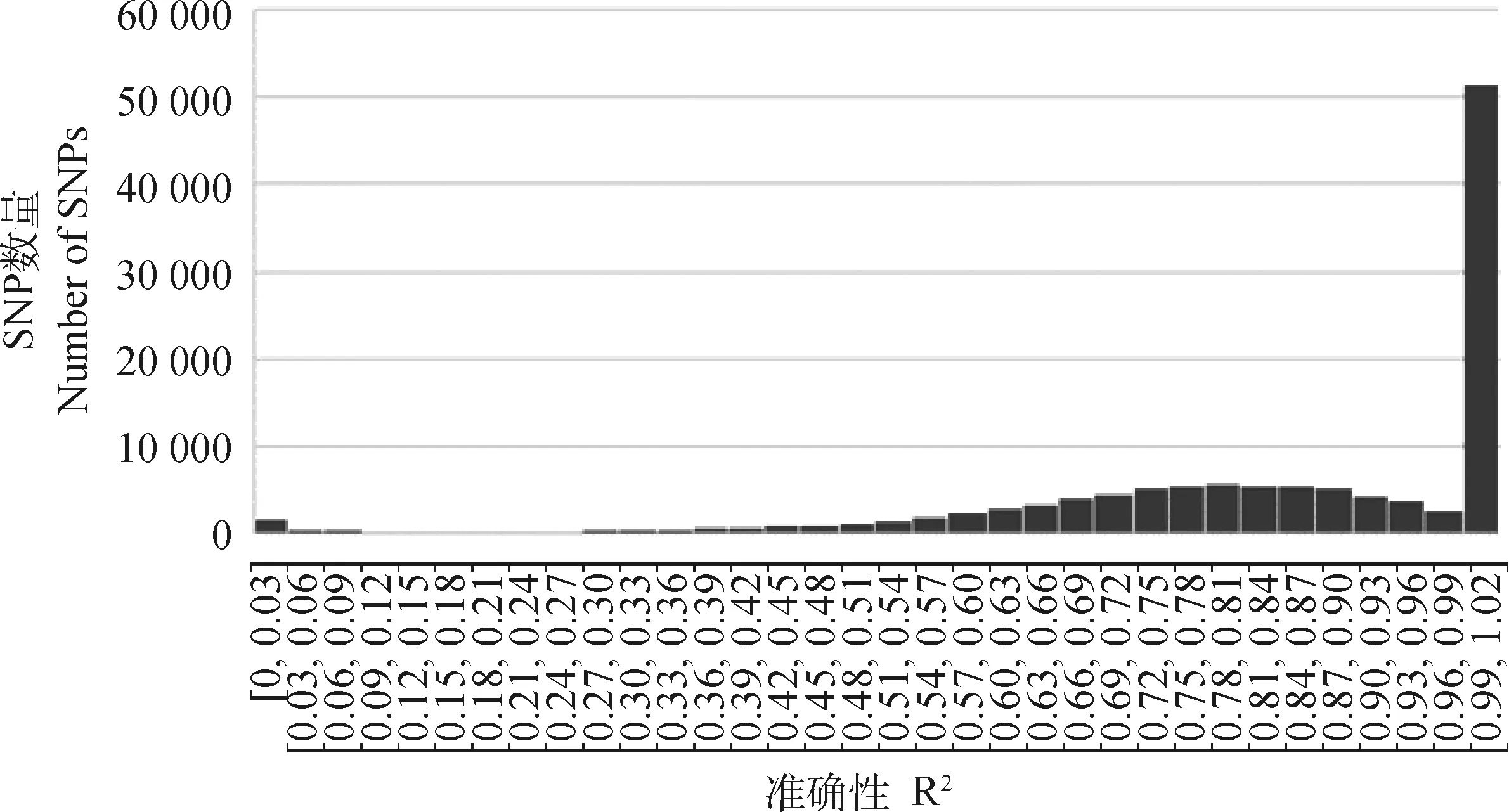
图8 填充准确性分布图

表4 不同组估计育种值
3 讨 论
自2008年启动奶牛群体遗传改良计划以来,我国奶牛群遗传质量明显提升,DHI测定群成年母牛平均单产水平逼近10吨。在新一轮的种业振兴大背景下,良种是现代奶业生产和发展的基础,而引进或者自产胚胎移植是培育种公牛和快速扩充生产群非常有效的技术手段[19-20]。因此,胚胎的质量直接决定着核心种源自主培育和奶牛良种快速扩繁的进程和速度。当下,人类胚胎植入前检测技术已经发展到了较高的水平,可以实现无创检测,筛选优质胚胎进行移植以保证受胎率和后代质量[16-17,21-22]。但是,牛胚胎遗传价值或生产性能的测定还处于起步阶段,相应的取样技术、微量DNA扩增和芯片检测等相关工作均未在国内开展。因此,本研究以体内、外胚胎为研究材料,探究利用胚胎微创取样技术对胚胎进行生产性能测定的可行性,以期为进口遗传物质监管和胚胎质量鉴定提供一套可行的操作方案。
近年来,随着活体采卵-体外受精技术在我国乃至世界范围的兴起[23-24],体外胚胎生产正在成为牧场优质种质资源扩繁的主要技术手段,但是体外胚胎受胎率和活犊率低仍然是当前产业发展面临的主要问题[5, 8]。如果能在植入前对胚胎进行质量鉴定,在排除一些自身存在染色体问题的胚胎基础上实现对胚胎生产性能的评估,那么将大大提高胚胎移植后的受胎率和犊牛的价值。因此,本研究通过胚胎切割技术对比分析了不同切割取样大小对取样后胚胎存活率、DNA扩增等方面的影响,结果表明利用胚胎切割仪切取少量滋养层细胞后,90%以上的胚胎可以继续发育孵化,提示这种取样方法适用于胚胎微创取样,这与Balvís等[25]的结果一致。
全基因组选择方法是通过检测芯片上SNP位点来评测个体的生产性能[26-27],为保证芯片SNP检测的准确性,芯片上机检测和数据分析前分别对DNA量和芯片SNP检出率都有明确规定,DNA量大于1 000 ng才能上机检测,芯片检出率高于90%才能进行芯片数据分析。微创取样的目的是尽可能减少对胚胎的损伤,但相应的问题就是细胞量少导致DNA初始含量过低,以至于无法应用常规的全基因组扩增方法进行扩增。为更均匀、准确的获得少量细胞的基因组结果,试验与中国农业大学高帅课题组合作,使用MALBAC技术对切取的少量TE进行扩增,MALBAC是线性放大[28],可以将获取的极少量胚胎细胞中的DNA均匀扩增上百万倍以满足基因分析的需求[29]。MALBAC扩增结果表明,各组扩增后DNA含量和OD值均符合要求,这其中1/2体内胚组和体外2细胞组DNA扩增成功率最高,体内滋养层组、体外滋养层组和8细胞组扩增成功率呈依次下降趋势。扩增后各组DNA量与扩增成功率趋势大体一致,起始细胞量越少,扩增后DNA量也越少。这说明在相同的扩增条件下,全基因组扩增成功率可能与起始DNA含量正相关,切割获取的起始DNA量越多,扩增成功率越高,与此同时取样细胞量与MALBAC扩增后DNA量也呈正相关。DNA扩增成功率与取样后胚胎存活率呈负相关,1/2体内胚组DNA扩增成功率和DNA量均最高,说明此扩增方法适用于牛的全基因组扩增,但是切取1/2胚胎对胚胎本身有较大损伤。为了减少切割造成的损伤,试验选择切取体内、外胚胎少量的滋养层细胞进行全基因组扩增,从DNA扩增量水平分析,两组扩增结果均符合芯片检测要求。
芯片检出率是芯片数据分析质控的另外一个关键环节[30-31],以100K芯片为例,当芯片检出率大于90%时,芯片检测数据可以进行后续的生产性能预测。考虑到MALBAC技术的特殊性,试验在胚胎样品之外又引入了荷斯坦奶牛全血样本的芯片数据作为阳性对照。与全血数据相比,胚胎各组芯片检出率只有1/2体内胚组无显著性差异,且检出率大于90%,其余各组均显著下降且检出率均低于90%,无法用于后续生产性能测定。基于MALBAC扩增的原理,推测这可能是因为起始细胞量太少导致的,在低起始量的情况下[29],扩增容易出现偏差和非特异性扩增。因此使用现有扩增条件,在保证胚胎正常发育的前提下,需要增加取样量。后续在取样技术没有突破之前,MALBAC技术的迭代尤其是针对牛的迭代更为重要。
试验进一步对检出率低于90%各组SNP缺失情况进行统计分析,结果显示体内、外滋养层组SNP缺失量比1/2体内胚组均高出了15倍以上,这说明微量取样后通过MALBAC扩增会导致更多的SNP缺失,而这些缺失的SNP是因为扩增导致还是因为本身发育所致需要进一步分析。为减少因全基因组扩增随机性导致的假片段缺失,试验扩大了染色体片段缺失所包含的连续SNPs数量,以SNP连续缺失7个(含7个)以上作为筛选标准,并对筛选到的缺失片段进行染色体定位分析和富集分析。染色体定位分析结果发现,各组染色体发生片段缺失的情况均不相同,其中chr1在两个试验组中均片段缺失量最多,这说明胚胎发育过程中可能存在真实的染色体片段缺失情况,进而导致了chr1在两个试验组中染色体片段缺失量明显上升。
胚胎工程技术的快速发展和广泛应用,极大提高了牧场中优良个体的繁殖力[32],但体外生产的胚胎常出现发育阻滞、胚胎质量差等问题[33-34],Raudsepp和Chowdhary[15]研究发现,染色体异常与胚胎质量、胚胎移植后妊娠率以及胎儿发育直接相关。染色体片段缺失是染色体异常较为常见的情况之一,缺失可能发生于任何一条染色体的任何位置,其发生原因情况较多,尤其在体外胚胎生产过程中,培养液、培养环境、人为操作等均可能对其造成影响。而且细胞骨架与细胞分裂时纺锤体形成、染色体运动、胞质分裂等事件密切相关[35-38]。因此,本研究比较了体内、外滋养层组的染色体缺失情况,并对缺失片段中包含的基因进行功能富集分析。结果发现,体外滋养层组排名前5的条目中有3个条目intermediate filament(GO:0005882)、keratin filament(GO:0045095)、intermediate filament cytoskeleton(GO:0045111)均与微丝蛋白和细胞骨架相关,共包括MARK2、SBNO2、GJA1等15个相关的基因(图9)。其中MARK2(microtubule affinity regulating kinase 2)编码的有丝分裂着丝粒相关蛋白能够调控有丝分裂微管的形成和长度[39],同时在间期和有丝分裂过程中,MARK2还能调控肌动蛋白和微管细胞骨架的变化,从而维持细胞的正常分裂[40]。值得注意的是,在体外滋养层组GO显著富集排名前30的条目中发还现十余个与细胞分化相关的条目,如osteoclast differentiation(GO:0030316)、positive regulation of cell differentiation(GO:0045597)等。众所周知,在胚胎发育过程中,调控细胞分化的任何一个环节出现问题,都有可能导致胚胎质量降低甚至直接导致胚胎发育异常[38]。本研究中,体外滋养层组染色体缺失片段中涉及到多个基因(如MYOD1、PRXL2A、FOXO3等)均可参与细胞分化(图9)。此外,体外滋养层组显著富集的KEGG通路中包含多条参与调控细胞分化的信号通路,如Hedgehog signaling pathway(bta04340)、cAMP signaling pathway(bta04024)、MAPK signaling pathway(bta04010)等。而体内滋养层组中发现的则更多的是调控细胞内离子通道等相关的通路,如p53 signaling pathway(bta04115)、Hippo signaling pathway(bta04390)、AMPK signaling pathway(bta04152)。这说明KEGG的富集结果与GO富集结果是相符的。鉴于在本研究中体外滋养层组的缺失基因显著富集到多个与细胞骨架和细胞分化相关的条目,推测体外胚质量差可能是由于胚胎在体外培养的过程中受不利因素影响导致调控细胞骨架相关的基因异常表达,从而直接导致染色体出现片段缺失、基因拷贝数异常等现象,进而影响胚胎发育过程中细胞分化的有序进行,最终影响胚胎的正常发育。
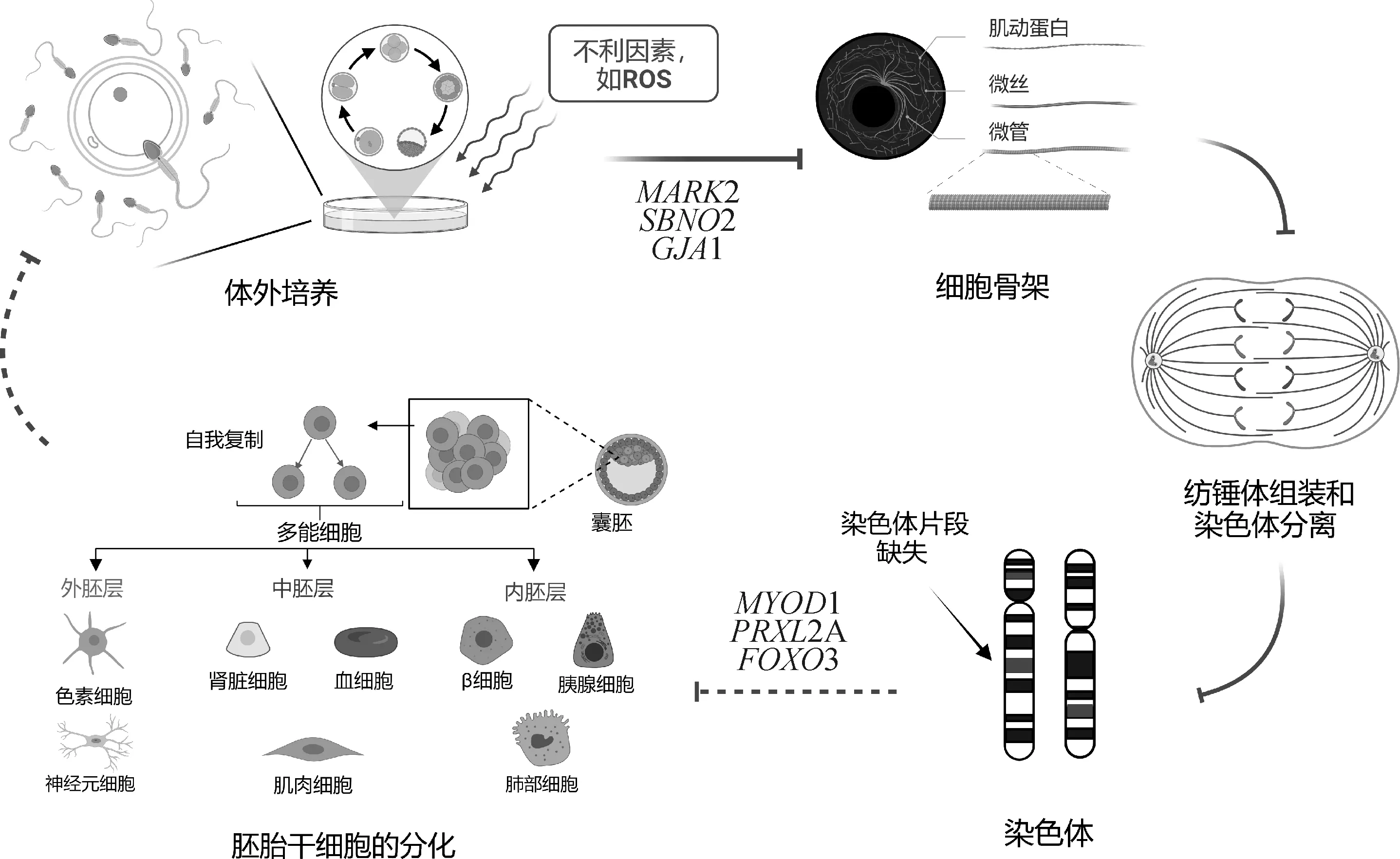
图9 体外滋养层组GO分析结果示意图
由于体内滋养层组和体外滋养层组检出率均小于90%,所以本研究对各组芯片数据进行了芯片填充,并估计其育种值。结果显示,本研究使用芯片包含95 257个SNPs位点,共填充64 958个SNPs位点,其中填充准确性在0.99~1之间的SNP位点有52 334 个。同时计算本次填充的准确性为91%,说明芯片填充效果较好,可用于育种值估计。同时,芯片填充后育种值估计能够很好的反映各组估计育种值的差异,以1/2体内胚组为例,样本PTP57001的产奶量、乳脂量和乳蛋白量的育种值都是最高的,从而可以判定胚胎PTP57001生产性能良好,可以用来移植。此外,从产奶量来看,体外滋养层组低于体内滋养层组,并且组内差异大,这可能是因为体内胚胎生产过程中所选的父母本通常生产性能良好,而屠宰场的母本一般生产性能低且水平不同,进而导致估计育种值低。
4 结 论
本研究结果表明,利用胚胎切割、单细胞基因组扩增和基因组芯片可在微创的前提下,实现对植入前早期胚胎染色体质量和生产性能的评估;此外,通过对比分析体内、外胚胎染色体片段缺失结果发现,胚胎体外发育过程中细胞骨架等基因异常表达可能是导致体外胚胎质量差的原因之一。
