LncRNA NEAT1通过调控miR-124-3p对食管鳞状细胞癌细胞迁移、侵袭的影响*
2022-11-21李险波张志强韩小勇甄志鹏
李险波,张志强,韩小勇,甄志鹏,赵 钦
保定市第一中心医院胸外科,河北保定 071000
食管癌是癌症死亡的第六大主要原因,也是世界第八大常见癌症类型[1]。食管癌多为食管鳞状细胞癌(ESCC),其占食管癌的90%以上,且侵袭性强,生物学行为复杂。ESCC主要由吸烟、饮酒或营养失衡等因素引起[2]。尽管外科手术、化学疗法或放射疗法等治疗方法改善了ESCC患者的预后,但是接受治疗的患者5年生存率仅为25%~30%[3-4]。因此,迫切需要探索用于治疗ESCC的分子靶标。
长非编码RNA(LncRNA)是一类长度大于200个核苷酸的RNA分子,不能编码蛋白质[5]。近年来,发现越来越多的LncRNA在肿瘤中差异表达,并且在肿瘤的发生、发展中起着至关重要的作用[6]。核富集转录本1(NEAT1)是一个约3.7 kb的核富集LncRNA,位于染色体11q13.1上[7]。相关研究表明,LncRNA NEAT1在膀胱癌、肺癌、乳腺癌和结直肠癌等多种恶性肿瘤中表达异常[8],且LncRNA可以充当天然“微小核糖核酸(miRNA)海绵”,阻止miRNA与信使RNA结合,抑制miRNA功能[9]。miR-124-3p作为近年来备受关注的miRNA之一,其在宫颈癌组织中低表达,且其表达水平与肿瘤浸润深度、组织分化程度、国际妇产科学联盟(FIGO)分期和淋巴结转移有关[10]。但LncRNA NEAT1及miR-124-3p在ESCC发生、发展中的具体生物学功能尚不清楚,因此本研究通过体外培养食管癌Eca109细胞,探讨LncRNA NEAT1及miR-124-3p对ESCC细胞迁移、侵袭的影响以及可能的分子机制,以期为ESCC的临床治疗提供重要的参考依据。
1 材料与方法
1.1细胞来源 ESCC细胞Eca109及正常人食管上皮细胞(HEEC)均购自美国典型培养物保藏中心(ATCC)。
1.2主要试剂与仪器 miR-124-3p模拟物(miR-124-3p mimics)及其阴性对照(miR-NC)、miR-124-3p抑制物(anti-miR-124-3p)及其阴性对照(anti-miR-NC)、NEAT1过表达物(OE-NEAT1)及其阴性对照(OE-NC)、LncRNA NEAT1小干扰RNA(si-NEAT1)及其阴性对照(si-NC)均购自美国ThermoFisher公司;RPMI 1640培养基、DMEM培养基、胎牛血清(FBS)均购自TaKaRa公司;Transwell小室购自金图生物科技有限公司;Lipofectamine 2000转染试剂盒购自美国Bio-Rad公司;TaqMan反转录试剂盒、TransStart®Green qPCR SuperMix、双荧光素酶报告基因检测试剂盒均购自北京全式金生物科技有限公司;荧光定量PCR仪、CO2培养箱均购自德国Sigma公司。
1.3方法
1.3.1细胞培养与转染 将HEEC、Eca109细胞均置于含10%FBS的DMEM培养基中,在37 ℃、5%CO2的培养箱中进行常规原代培养。收集对数期生长的Eca109细胞,以1×106个/孔的密度接种到6孔培养板中,待细胞融合率达到80%左右时,利用Lipofectamine 2000转染试剂盒对细胞进行分组转染。细胞分为空白组(细胞未转染)、miR-NC组(细胞转染50 pmol/μL miR-NC)、miR-124-3p mimics组(细胞转染50 pmol/μL miR-124-3p mimics)、anti-miR-NC组(细胞转染50 pmol/μL anti-miR-NC)、anti-miR-124-3p组(细胞转染50 pmol/μL anti-miR-124-3p)、OE-NC组(细胞转染5 μL OE-NC)、OE-NEAT1组(细胞转染5 μL OE-NEAT1)、si-NC组(细胞转染5 μL si-NC)、si-NEAT1组(细胞转染5 μL si-NEAT1)、anti-miR-124-3p+si-NC组(细胞共转染50 pmol/μL anti-miR-124-3p和5 μL si-NC)、anti-miR-124-3p+si-NEAT1组(细胞共转染50 pmol/μL anti-miR-124-3p和5 μL si-NEAT1),转染48h 后,用于后续实验。
1.3.2实时荧光定量PCR(qPCR)检测LncRNA NEAT1及miR-124-3p表达水平 使用Trizol试剂进行总RNA提取,利用TaqMan反转录试剂盒将总RNA逆转录为cDNA。使用TransStart®Green qPCR SuperMix试剂盒对LncRNA NEAT1及miR-124-3p进行定量。扩增反应体系:2×TransStart®Green qPCR SuperMix 10 μL,正向引物(10 μmol/L)0.5 μL,反向引物(10 μmol/L)0.5 μL,cDNA模板(1 μg)2 μL,使用RNase-Free H2O补足至20 μL。扩增条件:95 ℃,30 s;95 ℃,5 s;60 ℃,20 s;72 ℃,10 s,共40个循环。以GAPDH为内参,标准化LncRNA NEAT1值;以U6为内参,标准化miR-124-3p值。最后采用2-ΔΔCt法计算LncRNA NEAT1、miR-124-3p表达水平。所用引物序列见表1。

表1 LncRNA NEAT1、miR-124-3p、GAPDH、U6引物序列
1.3.3Transwell实验检测Eca109细胞迁移情况 收集各组Eca109细胞,将细胞水平调整为1×104/mL,将Eca109细胞接种到100 μL无血清的RPMI 1640培养基中,然后加入到Transwell上腔室中。同时,将500 μL含10%FBS的RPMI 1640培养基添加到下腔室。在37 ℃下孵育24 h后,将细胞用0.1%结晶紫染色20 min,最后在光学显微镜下计数迁移的细胞数目。
1.3.4Transwell实验检测Eca109细胞侵袭情况 在上腔室中预先涂上Matrigel,37 ℃干燥1 h。收集各组Eca109细胞,将细胞水平调整为1×104/mL,其余步骤同上述1.3.3。
1.3.5双荧光素酶报告基因实验验证LncRNA NEAT1与miR-124-3p靶向关系 使用StarBase v3数据库预测LncRNA NEAT1与miR-124-3p结合位点。分别构建LncRNA NEAT1的野生型(WT)和突变型(MUT)3′-UTR区质粒,标记为WT-NEAT1、MUT-NEAT1。将Eca109细胞置于含10%FBS的DMEM培养基中,于37 ℃,5%CO2培养箱里培养,待Eca109细胞密度达到80%左右时,按照miR-NC+WT-NEAT1共转染组、miR-124-3p mimics+WT-NEAT1共转染组、miR-NC+MUT-NEAT1共转染组、miR-124-3p mimics+MUT-NEAT1共转染组转染Eca109细胞。转染48 h后,通过双荧光素酶测定系统进行荧光素酶报告基因测定。

2 结 果
2.1HEEC、Eca109细胞中LncRNA NEAT1、miR-124-3p表达水平比较 与HEEC细胞相比,Eca109细胞中LncRNA NEAT1表达水平明显升高,而miR-124-3p表达水平明显降低,差异均有统计学意义(P<0.05),见图1。
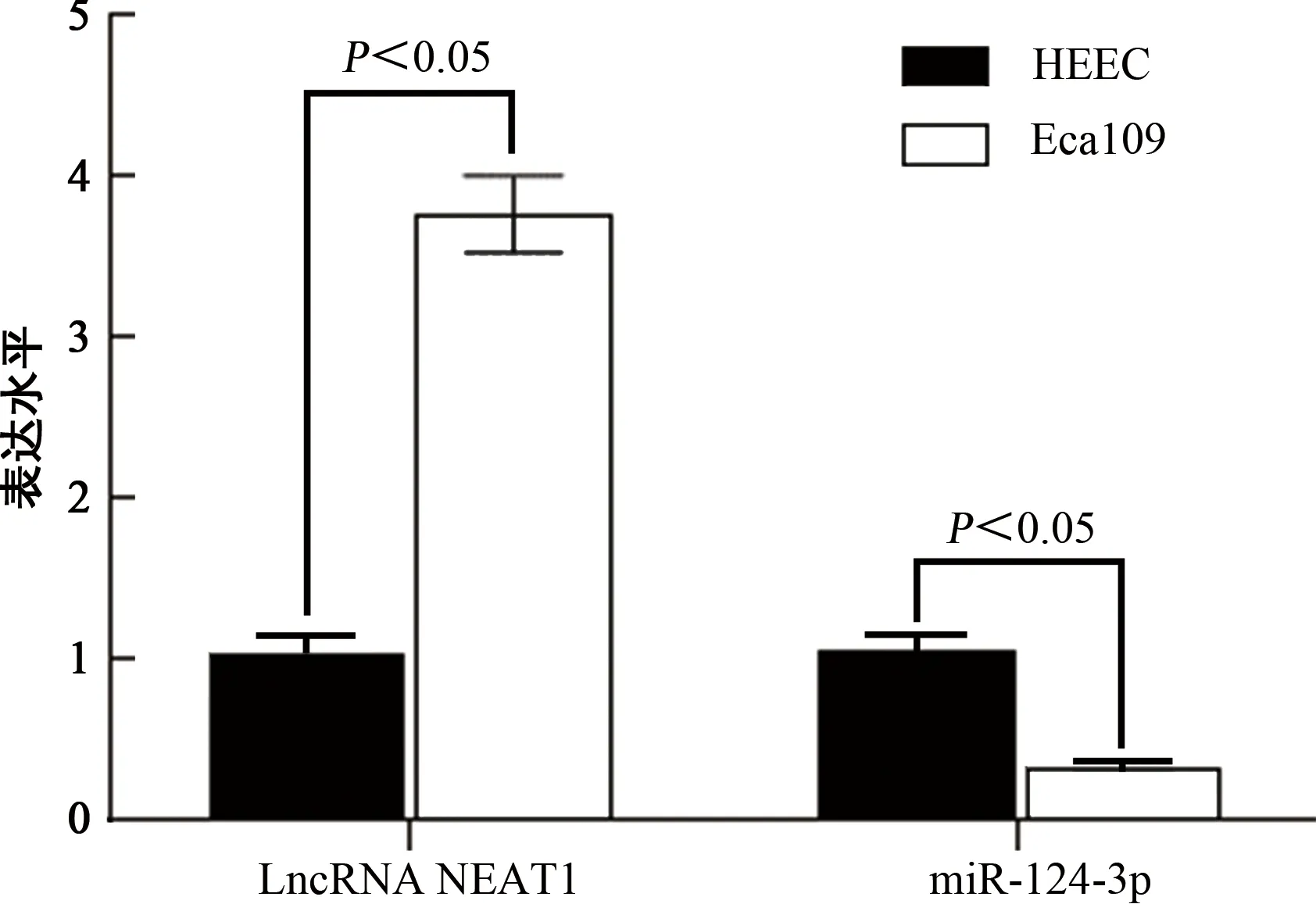
图1 HEEC和Eca109细胞中LncRNA NEAT1、miR-124-3p表达水平比较
2.2下调LncRNA NEAT1表达水平对Eca109细胞迁移、侵袭的影响 qPCR结果显示, si-NEAT1组Eca109细胞中LncRNA NEAT1表达水平明显低于空白组和si-NC组(P<0.05),提示Eca109细胞转染si-NEAT1成功,见图2A。Transwell实验结果显示,与空白组和si-NC组比较,si-NEAT1组Eca109细胞迁移、侵袭数均明显降低(P<0.05),见图2B、C、D、E。
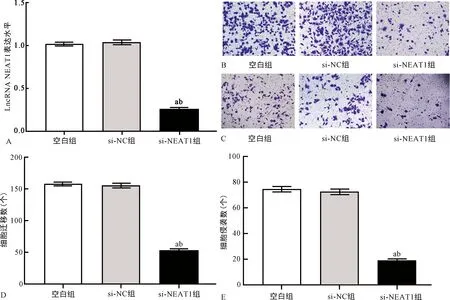
注:A为空白组、si-NC组、si-NEAT1组Eca109细胞中LncRNA NEAT1表达水平比较;B、C为光学显微镜下(×400)空白组、si-NC组、si-NEAT1组Eca109细胞迁移(B)、侵袭(C)结果;D、E为空白组、si-NC组、si-NEAT1组Eca109细胞迁移(D)、侵袭(E)数比较;与空白组比较,aP<0.05;与si-NC组比较,bP<0.05。图2 下调LncRNA NEAT1表达水平对Eca109细胞LncRNA NEAT1表达水平及迁移、侵袭能力的影响
2.3miR-124-3p过表达对Eca109细胞迁移、侵袭的影响 miR-124-3p mimics组Eca109细胞中miR-124-3p表达水平明显高于空白组和miR-NC组(P<0.05),提示Eca109细胞转染miR-124-3p mimics成功,见图3A。与空白组和miR-NC组比较,miR-124-3p mimics组Eca109细胞的迁移、侵袭数均明显降低(P<0.05),见图3B、C、D、E。

注:A为空白组、miR-NC组、miR-124-3p mimics组Eca109细胞中miR-124-3p表达水平比较;B、C为光学显微镜下(×400)为空白组、miR-NC组、miR-124-3p mimics组Eca109细胞迁移(B)、侵袭(C)结果;D、E为空白组、miR-NC组、miR-124-3p mimics组Eca109细胞迁移(D)、侵袭(E)数比较;与空白组比较,aP<0.05;与miR-NC组比较,bP<0.05。图3 miR-124-3p过表达对Eca109细胞LncRNA NEAT1表达水平及迁移、侵袭能力的影响
2.4LncRNA NEAT1靶向调控miR-124-3p的表达 利用StarBase v3数据库预测发现LncRNA NEAT1与miR-124-3p存在结合位点,见图4A。共转染双荧光素酶报告基因实验结果显示,miR-124-3p mimics+WT-NEAT1组Eca109细胞的荧光素酶相对活性较miR-NC+WT-NEAT1组明显降低(P<0.05),而miR-124-3p imics+MUT-NEAT1组、miR-NC+MUT-NEAT1组Eca109细胞荧光素酶相对活性差异无统计学意义(P>0.05),见图4B。qPCR检测结果显示,si-NEAT1组Eca109细胞中miR-124-3p表达水平明显高于空白组和si-NC组(P<0.05),而OE-NEAT1组miR-124-3p表达水平明显低于空白组和OE-NC组(P<0.05),见图4C。
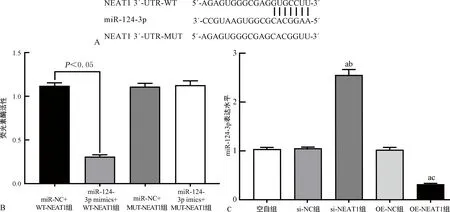
注:A为StarBase v3数据库预测LncRNA NEAT1与miR-124-3p的结合位点;B为双荧光素酶报告基因实验结果;C为LncRNA NEAT1下调和过表达对miR-124-3p的靶向调控作用;与空白组比较,aP<0.05;与si-NC组比较,bP<0.05;与OE-NC组比较,cP<0.05。图4 LncRNA NEAT1靶向调控miR-124-3p的表达
2.5抑制miR-124-3p表达逆转了下调NEAT1表达水平对Eca109细胞迁移、侵袭的作用 qPCR结果显示,si-NEAT1+anti-miR-124-3p组Eca109细胞中miR-124-3p表达水平明显低于si-NEAT1组和si-NEAT1+anti-miR-NC组,提示Eca109细胞共转染anti-miR-124-3p和si-NEAT1成功,见图5A。与si-NEAT1组和si-NEAT1+anti-miR-NC组比较,si-NEAT1+anti-miR-124-3p组Eca109细胞的迁移、侵袭数均明显升高(P<0.05),见图5B、C、D、E。

注:A为si-NEAT1组、si-NEAT1+anti-miR-NC组、si-NEAT1+anti-miR-124-3p组Eca109细胞中miR-124-3p表达水平比较;B、C为光学显微镜下(×400)si-NEAT1组、si-NEAT1+anti-miR-NC组、si-NEAT1+anti-miR-124-3p组Eca109细胞迁移(B)、侵袭(C)结果;D、E为si-NEAT1组、si-NEAT1+anti-miR-NC组、si-NEAT1+anti-miR-124-3p组Eca109细胞迁移(D)、侵袭(E)数比较;与si-NEAT1组比较,aP<0.05;与si-NEAT1+anti-miR-NC组比较,bP<0.05。图5 同时抑制miR-124-3p及LncRNA NEAT1表达对Eca109细胞迁移、侵袭能力的影响
3 讨 论
肿瘤细胞的迁移、侵袭被认为是恶性肿瘤最重要的生物学特性之一,是恶性肿瘤发生、发展的重要因素。据统计,约90%的癌症患者死于肿瘤细胞侵袭和迁移[11]。ESCC是常见的癌症类型之一,由于缺乏特异性症状和有效的早期诊断方法,导致大多数ESCC患者确诊时已出现肿瘤局部浸润和远处转移,随之而来的是肿瘤浸润、转移引起的并发症,这成为ESCC患者死亡的主要原因[12-13]。因此,更好地了解ESCC肿瘤发生的分子机制和筛选生物标志物,对ESCC的早期诊断和治疗具有重要意义。
近年来,研究发现LncRNA通过表观遗传修饰、转录调控、miRNA海绵等多种途径驱动肿瘤的发生与发展[14],差异表达的LncRNA与肿瘤的增殖、侵袭、转移和预后密切相关,有可能成为肿瘤治疗的新靶点[15]。据报道,LncRNA NEAT1在喉鳞状细胞癌组织中的表达水平明显高于癌旁组织,且颈淋巴结转移或临床分期晚期患者的LncRNA NEAT1表达水平较高[16];LncRNA NEAT1在ESCC细胞中高表达,能增强ESCC细胞活力和侵袭能力[17];沉默LncRNA NEAT1后,胃癌细胞的增殖及侵袭能力明显降低[18]。表明LncRNA NEAT1在多种肿瘤中具有促癌作用。本研究结果显示,LncRNA NEAT1在ESCC细胞中高表达,下调LncRNA NEAT1表达水平可明显抑制Eca109细胞的迁移、侵袭能力,这与前述研究结果一致。提示LncRNA NEAT1在ESCC中发挥癌基因的作用,LncRNA NEAT1可能成为治疗ESCC的潜在靶点。
LncRNA可作为一种竞争性内源性RNA与miRNA相互作用,参与靶基因的表达调控[19]。为了进一步探究LncRNA NEAT1在ESCC细胞迁移、侵袭中的具体分子机制,本研究通过StarBase数据库预测LncRNA NEAT1与miR-124-3p可以靶向结合,并通过双荧光素酶报告基因实验证实miR-124-3p是LncRNA NEAT1的靶基因。据相关研究报道,miR-124-3p能抑制肝癌增殖、迁移、侵袭和淋巴道转移,同时具有促进肝癌细胞凋亡的作用[20];miR-124-3p可通过下调靶基因STAT3的表达,进而影响其下游的p-STAT3、细胞周期素D2和基质金属蛋白酶2信号通路,从而促进鼻咽癌细胞凋亡,抑制鼻咽癌细胞的增殖、迁移及侵袭[21];miR-124-3p还能通过靶向CRKL抑制肝癌细胞的迁移、侵袭[22];miR-124-3p也可通过靶向整合素β3抑制胃癌细胞的迁移、侵袭,且miR-124-3p可能成为治疗胃癌的分子靶标[23]。本研究结果显示,miR-124-3p过表达可明显抑制Eca109细胞迁移、侵袭,提示miR-124-3p在ESCC中发挥着抑癌作用。为进一步验证LncRNA NEAT1靶向调控miR-124-3p对ESCC细胞的迁移和侵袭作用,本研究将Eca109细胞共转染si-NEAT1和anti-miR-124-3p,结果发现,抑制miR-124-3p表达逆转了下调LncRNA NEAT1表达水平对Eca109细胞迁移、侵袭的抑制作用。提示下调LncRNA NEAT1表达水平对ESCC细胞的迁移、侵袭涉及的分子机制可能与激活miR-124-3p表达有关。LIU等[24]的研究表明LncRNA NEAT1敲低通过上调miR-124-3p表达水平而减弱肝癌细胞生长,与本研究结果一致。本研究只是在体外细胞水平上证明了LncRNA NEAT1/miR-124-3p在ESCC细胞迁移、侵袭过程中的作用,后续体内实验尚需进一步深入研究。
综上所述,下调LncRNA NEAT1表达水平可能通过激活miR-124-3p的表达来抑制ESCC细胞的迁移、侵袭能力,其为ESCC生物靶向治疗提供了可能的靶点。
