双荧光素酶报告基因系统在家蚕基因启动子研究中的应用
2021-03-18欧阳萃鸿陈太生谢晓乐李荣松
欧阳萃鸿 陈太生 谢晓乐 李荣松 田 铃
(华南农业大学动物科学学院 广东广州 510642)
启动子是基因结构中一段可以被RNA 聚合酶识别、结合、起始转录的DNA 序列,它含有RNA 聚合酶特异性结合和转录起始所需的保守序列。启动子本身不被转录,而是通过其中的反应元件(response elements)与转录因子(transcription factor)结合进而控制基因的转录活动。转录调控是调节基因功能最直接、有效的方式[1]。将目的基因的启动子区域构建至双荧光素酶报告基因表达载体中,通过检测双荧光素酶报告基因系统荧光蛋白的酶活,从而分析目的基因启动子的转录活性,这是基因转录调控研究中常用且有效的手段之一[2]。家蚕是鳞翅目昆虫的模式生物,本文以家蚕基因启动子调控机制的研究为切入点,对双荧光素酶报告基因系统的工作原理与特点、研究和应用现状进行综述,有助于我们从细胞和分子层面理解昆虫生长发育过程的基因转录表达调控机制,为今后该系统在昆虫中的深入应用提供思考。
1 双荧光素酶报告基因系统的发展历史
荧光素酶是最初从自然界发光生物中获得的一种可以催化对应的荧光底物发生氧化反应产生荧光的蛋白酶类物质,哺乳细胞没有内源性荧光素酶[3]。荧光底物与酶发生反应产生的光其本质是一种生物萤光,属于可见光,不依赖外源激发光作用,在自然界一些发光生物身上通常能见到这种生物萤光。而荧光则是通过激发光照射荧光物质,光能使荧光物质原子核周围的电子发生跃迁反应,由原来的低能量电子轨道跃迁到高能量电子轨道,但是这种原子状态并不稳定,即原子会从激发态回到基态,原先电子从激发光中获得的能量又会重新以光能的形式释放出去,形成具有一定的波长特征的荧光。所以荧光和萤光是有一定区别的,Promega 公司为了区分这两类光信号的不同一直将Luciferase 称为萤光素酶,但惯例上我们还是称其为荧光素酶。
地球存在许多发光的生物,如萤火虫等。在1884年,法国科学家Dubois 经实验认为,荧光素底物、荧光催化酶以及氧是发光反应的基本条件,由此开始了对萤火虫发光现象的研究与应用。随后的数十年内,科学家们不断地对萤火虫发光物质本质进行探索和查找。1942年,美国约翰霍普金斯大学麦科勒-普拉特研究所(The McCollum-Pratt Institute and the Department of Biology, Johns Hopkins University)的McElory 发现三磷酸腺苷(adenosine triphosphate, ATP)直接参与了萤火虫发光反应[4]。1954年,McElory 等人从发光的细菌中提取到了荧光素酶结晶,由此发光生物荧光素酶的研究迈上了新的台阶[5]。随后在 1956年,Green 与McElory 等一起第一次从萤火虫中提取得到了较高纯度的萤火虫荧光素酶结晶制剂,自此开始了萤火虫荧光素酶在生物学研究中的开发和应用[6]。
除萤火虫类昆虫外,目前在自然界中已经发现了多种可以发光的生物,人们对这些来源于不同发光生物的荧光素酶进行了提取,并分类为萤火虫荧光素酶(firefly luciferase)、细菌荧光素酶(bacterial luciferase)和另外一些从发光甲虫、发光海洋生物提取的荧光素酶[3]。萤火虫荧光素酶最早提取于一种北美萤火虫(Photinus pyralis),目前已在世界各地的萤火虫中提取到萤火虫荧光素酶[3]。细菌荧光素酶是从明亮发光杆菌(Photobacterium phosphoreum)、羽田希瓦氏菌(shezoanella hanedai)、夏威夷弧菌(Vibrio harvey)、青海弧菌(V.qinhaiensis)、火神弧菌(V.logei)、费氏弧菌(V.fischeri)等多种海洋和淡水中的发光菌中提取获得的。在1960年,美国乔治亚大学(University of Georgia)的Cormier 等研究者从海肾(renilla)中提取了海肾荧光素酶(Renilla reniformisluciferase),为海肾荧光素酶在科研中的应用打下了基础[7]。
数十年来,研究者们一直尝试将荧光素酶作为一种报告基因来研究基因启动子的活性和蛋白质之间的相互作用。早在1988年就有通过对荧光素酶报告基因表达的蛋白的荧光强度进行检测,进而综合分析不同处理条件对基因启动子活性影响[8]。这就是典型的、利用荧光素酶报告基因系统表达产物的特性建立起来的一种可以对目的基因转录活性进行定量检测的技术体系[3]。在1990年,经过两年的研究与发展形成了一套相对成熟的荧光素酶生物传感器技术,同年其发明者加入了Promega 公司。经过几年的潜心研发,1996年Promega 公司正式发布了双荧光素酶报告基因系统并开始销售相关检测试剂,这种技术创造性地结合了萤火虫荧光素酶和海洋腔肠荧光素酶两种荧光检测体系,是荧光素酶技术在漫长发展历史中的一次重要的突破。双荧光素酶报告基因系统与传统共报告基因系统相比较,具有更简便、高效、快速的特点,因此在分子生物学诸多领域的研究中得到了应用。
2 双荧光素酶报告基因系统的基本工作原理
理想的报告基因应该具有以下几个基本特点:①具有已知且可被克隆验证的全部基因序列;②报告基因的表达产物在检测中要反应灵敏,酶活反应迅速;③不与目的研究细胞中的任何表达基因相似,否则会干扰转染进入细胞内的荧光素酶的作用效果[9];④报告基因的表达产物有较宽的动态检测范围,具有良好的稳定性;⑤报告基因的表达产物不影响目的研究细胞的正常生理活动;⑥报告基因在目的研究细胞中的表达产物易于检测,且能够按照标准读取量值。大部分的报告基因表达产物通常是酶,可以通过催化反应级联放大信号,再以萤光或放射性信号被检测[10]。
在几种荧光素酶中,萤火虫的荧光素酶最适合我们开展实验研究使用的。大多数细菌的荧光素酶最适的活性温度在15 ℃~25 ℃,温度超过25 ℃时其酶活会迅速下降,37 ℃下基本全部失活,所以不适合在动物细胞中使用[11]。相较而言,萤火虫荧光素酶则对催化温度要求较低,一般室温即可,适应温度范围广,因此该酶报告基因常用于各类真核细胞基因启动子活性的分析与检测。目前,有些新型荧光素酶如Gaussia 荧光素酶和Cypridina 荧光素酶,它们的基因携带了一种分泌信号肽,表达产物能分泌到细胞外,可以在不裂解破碎细胞的条件下直接检测酶活,甚至还能连续检测时间曲线上报告基因表达产物的变化,进而分析报告基因的表达活性变化,此技术方法适用于高通量筛选[12]。
萤火虫荧光素酶是一种可以催化荧光底物发生反应的单亚基特异活性蛋白,分子量为60~64 kDa。萤火虫荧光素酶报告基因的优点在于只要完成转录翻译,其表达产物就立刻具有可被检测的酶活[3]。在 ATP、二价镁离子(Mg2+)、氧气(O2)等存在的情况下,萤火虫荧光素酶会催使底物萤火虫荧光素发生氧化,消耗 ATP 和 O2产生Lucifery1-AMP (adenosine monophosphate,AMP)中间体(见图1),氧化的荧光素会从激发态回到基态释放出光子,产生波长540~600 nm 的黄绿色荧光信号,这类荧光信号可以通过发光检测仪或CCD(Charge-coupled Device)相机来进行检测或捕获[13]。但这一生化反应过程比较缓慢,因此在荧光素酶和荧光素底物混合后产生的荧光会迅速衰减[3],所以在专业化的试剂盒中会通过添加辅酶A(coenzyme A,CoA)来提高萤火虫荧光素酶的活性,加快该荧光反应的速度,解决荧光衰减的问题,产生连续的荧光信号。除辅酶A外,牛血清蛋白和氨基乙醇等物质也具有相同的作用[14]。

图1 萤火虫荧光素酶化学反应[15]
提取自海洋腔肠动物海肾的荧光素酶也是一种可以催化荧光素发生荧光反应的单亚基特异活性蛋白,其分子量为36 kDa。同萤火虫荧光素酶一样,该蛋白质在完成转录翻译后即具有催化活性[15]。海肾荧光素酶不依赖ATP 参与酶促反应,在有荧光底物腔肠素(coelenterazine)和O2等反应物质的条件下,海肾荧光素酶即可催化荧光底物腔肠素发生氧化,在反应过程会产生波长为460~540 nm 的蓝色荧光信号(见图2)。和萤火虫荧光素酶相似,海肾荧光素酶反应产生的荧光信号也会在短时间内迅速衰减。这种报告基因既可以作为内参与萤火虫荧光素酶等其他报告基因一起组成双检测系统,也可以作为一种单独的检测系统用于实验。天然腔肠素在适宜条件下能够穿越活细胞的细胞膜,与活细胞内表达的海肾荧光素酶发生酶促反应产生荧光,因此海肾荧光素酶报告基因适用于有活细胞检测需求的实验[11]。
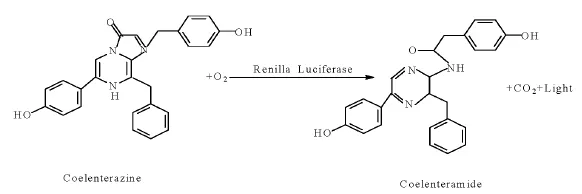
图2 海肾荧光素酶化学反应[15]
Promega 公司是最早将萤火虫荧光素与海洋腔肠荧光素酶两种不同荧光检测体系相结合提供双荧光素报告基因系统的生物制剂公司,该检测系统是一种含有内参、可归一化处理实验数据、分析实验现象的方案[16]。萤火虫荧光素酶和海洋腔肠荧光素酶检测到的是具有不同特征的数据,归一化处理可以使这些数据变得具有可比性的同时又保留一定的相对关系。即在实验中可以通过设置合理的内参,让实验预处理过程得到的没有可比性的数据得到一定的限制,能够对照参照物进行计算,进而消除各种实验差异如细胞数、转染效率、细胞状态、裂解效率、检测温度、时间等因素带来的影响[17]。
早期的双荧光素酶报告系统需要裂解细胞,并先后完成两个生物荧光反应的定量检测,再对实验数据进行处理和分析,以达到对两个同时表达的报告基因的表达活性进行检测的目的[18]。实验步骤是首先将构建好的、带有目的基因启动子的报告基因载体和内参载体共转染到培养细胞中进行表达,检测荧光时要用裂解液将转染的细胞进行破碎裂解,然后加入萤火虫荧光素与萤火虫荧光素酶发生荧光反应。随后,用荧光检测工具对荧光强度进行测定,检测完成后还需淬灭该荧光反应,再加入腔肠素与海肾荧光素酶发生荧光反应,检测其荧光强度[17]。其中萤火虫荧光素酶作为主报告基因,其表达水平随外界的处理而发生变化,海肾荧光素酶作为内参报告基因,其表达水平一般不会随外界处理而发生变化[18]。
目前,Thermo 公司研发了一种比较便捷的双分泌型萤光素酶报告基因检测技术,其选用的是Gaussia 分泌型荧光素酶和Cypridina 分泌型荧光素酶[19]。这两种酶均为分泌蛋白,在表达过程中会通过内质网分泌到细胞外,因此实验时不需要裂解细胞,取培养液上清就能进行荧光检测。但是这两种酶的荧光信号波长相似,一起检测的话难以分辨,一般要将测试样品分为相同的两组再分别用不同的荧光素底物对酶活性进行检测,借此进行区分。这种检测技术不用考虑荧光素酶在细胞内底物的影响,比萤火虫荧光素酶有更高的检测灵敏度和更广的动态检测范围[20]。
3 双荧光素酶报告基因系统在家蚕基因启动子研究中的应用
在1995年,国内就有研究者首先将野生型家蚕BmNPV病毒基因组DNA 与克隆带有荧光素酶基因的pBmL 质粒进行重组,再将重组后的载体转染到家蚕细胞中进行表达,获得载有荧光素酶基因的重组病毒rBmNPVLu。研究发现,该重组病毒带有的荧光素酶基因能够在家蚕细胞中进行转录表达。随后,将重组病毒注射进家蚕幼虫体内,检测发现与其他病毒基因一样,荧光素酶基因在家蚕幼虫和蛹中注射重组病毒后的第5 天左右表达最高[21]。
3.1 相似性基因启动子活性差异的研究
家蚕有3 个BmSDH(Bombyx mori sorbitol dehydrogenase,BmSDH)基因,即BmSDH-1、BmSDH-2a、BmSDH-2b。其中BmSDH-2a和BmSDH-2b在基因结构和蛋白结构上都具有很高的相似性,二者可能具有相似的生理功能,而BmSDH-1则存在较大差异。于是朱娟等人利用PCR 技术克隆这3 个基因的启动子,分别构建带有萤火虫荧光素酶报告基因的载体,并与pRL-CMV报告质粒(含海肾荧光素酶报告基因)共转染家蚕BmN 细胞,通过双荧光素酶检测系统检测BmSDH基因启动子的活性。实验结果表明,BmSDH-2a的启动子活性极显著高于BmSDH-1和BmSDH-2b[22]。谢雨辰等人推测,家蚕卵内山梨醇脱氢酶的活性主要来自BmSDH-2a基因的表达。他们利用滞育激素、保幼激素(juvenile hormone,JH)和蜕皮激素(20 -hydroxyecdysone,20 E)处理转染有BmSDH-2a基因启动子(序列长度1082 bp)的荧光素酶报告基因系统的家蚕细胞,发现滞育激素不能调节BmSDH基因的表达,而高浓度的保幼激素和蜕皮激素则会抑制BmSDH基因的表达[22]。
赵斯斯等人发现家蚕细胞色素P450 家族CYP9A19和CYP9A22两个基因的表达存在组织特异性,CYP9A19在中肠、丝腺、脂肪体、马氏管中均有较高水平的表达;CYP9A22在中肠、丝腺、马氏管中有表达,但在脂肪体中无表达。为了研究这两2 个基因的表达调控机制,其构建了含荧光素酶报告基因和启动子不同长度顺次缺失片段的重组质粒,采用双荧光素酶报告基因检测系统检测启动子活性。结果表明:家蚕CYP9A19基因起始密码子游-1492~-1102 bp区域是重要的转录活性区域;CYP9A22基因启动子起始密码子上游-1630~-1210 bp 这一区域是重要的转录活性区域。用不同浓度的氟化钠诱导细胞后,发现0.1×10-3mol/L的NaF 能使荧光素酶活性增强显著,即0.1×10-3mol/L NaF能上调这2 个基因的表达[15]。
3.2 基因启动子序列区域调控元件的研究分析
谢雨辰等通过双荧光素酶报告基因系统对BmSDH-2a基因启动子反应元件进行鉴定,结果显示在BmSDH-2a启动子序列中-355~-674 bp 和-674~-1082 bp 的两个区域之间存在负调控元件[23]。庄兰芳等利用双荧光素酶报告基因系统对家蚕热激蛋白hsp70基因5’端的序列:hsp70 -1~hsp70 -305 和hsp70 -1~hsp70 -538 进行比较,结果显示片段hsp70 -1~hsp70 -538 的表达活性是hsp70 -1~hsp70-305 的1~3 倍,并且hsp70 -1~hsp70 -538 除了包含共有的热激元件(heat shock element,HSE)外,还包含3 个可能的HSE 元件[24]。
Li 等研究者以家蚕为模型,利用双荧光素酶报告基因系统证明了20 E 信号传导的初级应答基因E75不同亚型介导的蜕皮甾类生物合成与20 E 信号传导之间存在的调节环路。结果显示,E75基因的亚型A 和C 可以直接与Halloween基因启动子区域中的视黄酸受体相关反应元件结合,以诱导基因表达,从而促进蜕皮甾类生物合成和发育提前,而亚型 B 通过物理相互作用拮抗亚型A/C 对Halloween基因的转录活性。由于20 E 差异诱导E75基因亚型的表达,因此E75基因介导的调节环代表了类固醇生成的良好自动调节,这有助于精确控制发育时机[25]。
细胞自噬的发生需要在多个细胞自噬相关(autophagyrelated, Atg)蛋白的参与下才能完成[26]。Tian 等通过双荧光素酶报告系统和凝胶迁移等一系列实验研究发现,20 E 通过其受体EcR/Usp 结合到关键自噬基因Atg1的启动子区域反应元件诱导其表达,从而促进细胞自噬的发生[26]。顾健健等利用双荧光素酶报告系统在已报道的BmVgP78 M启动子上游利用基因工程连接了一段BrC-Z2 转录因子结合序列(BrC-Z2 element,BrC-Z2E),连接的这段序列可以对20 E进行应答并增强该启动子活性[27]。
陈恩祥研究发现,家蚕BmVgR(Bombyx mori vitellogenin receptor)基因的启动子均存在大量的POU 结构域,他们利用荧光素酶报告系统、凝胶阻滞实验(electrophoretic mobility shift assay, EMSA)和染色质免疫共沉淀(chromatin immunoprecipitation assay,ChIP)等实验验证了家蚕POU 结构域转录因子POU-M2 能够结合到BmVgR启动子上并启动BmVgR的高表达。同时,他们还研究发现20 E 能够诱导BmVgR的转录,20 E 信号传导因子BrC-Z1 能够结合到POU-M2基因的启动子区域诱导其表达,进而增强BmVgR的表达[28]。
3.3 研究小RNA 对靶向基因转录表达的影响
小RNA 为长度小于200 nt 的非编码RNA,主要包括微RNA(microRNA,miRNA)、小干扰RNA(small interfering RNA,siRNA)和Piwi-interacting RNA(piRNA)等,在转录和转录后水平上调控,以发挥基因沉默的作用。
家蚕细胞质多角体病毒(Bombyx moricytoplasmic polyhedrosis virus,BmCPV)感染家蚕后诱导的胰岛素相关肽结合蛋白2(Insulin-related peptide-binding protein2,IBP2)基因,可能是小分子RNA miR-278 -3P 的靶标之一。Wu 等构建PmirGLO-IBP2 和PmirGLO-IB2P2-mut(突变型)双荧光素酶报告基因载体,研究发现PmirGLO-IBP2和miR-278 -3p 模拟物共转染的细胞中,荧光素酶活性显着降低;在PmirGLO-IBP2-mut 和miR-278 -3p 模拟物共转染的细胞中,萤光素酶活性没有变化。这表明,在BmCPV感染后,可高度诱导B.mori IBP2基因的表达,并且被miR-278 -3p 负调控[29]。
根据家蚕核型多角体病毒(Bombyx morinucleo polyhedrosis virus,BmNPV)感染家蚕后小RNA 表达的变化,姜晓旭预测miR-277 -5p 可靶向作用于家蚕DNA 甲基转移酶基因Dnmt(DNA methyltransferase),通过构建PmirGLO-Dnmt2和PmirGLO-Dnmt2-mut(突变型)双荧光素酶报告基因载体等研究发现miR-277 -5p 对Dnmt基因表达具有显著的抑制作用[30]。
此外,双荧光素酶基因报告系统也可用于检测家蚕生长发育过程中小RNA 对基因转录表达的调控。刘晓等利用实时荧光定量PCR技术研究发现piR-4682(Piwi-interacting RNA-4682)在家蚕精巢、卵巢和血淋巴中表达量较高。随后,研究人员还发现在piR-4682 相同的作用位点存在另一个miRNA-305,它们均靶向调控small wing基因,将该基因的CDS(coding sequence)片段构建在psiCHECK 荧光素酶报告基因载体上,得到重组载体psiCHECK [small wing]。piR-4682 和miR-305 的类似物对small wing基因的表达都有一定抑制作用[31]。
陈恩祥等筛选出10 个从卵黄发生期到卵壳形成期上调表达的miRNAs 可能参与调控BmVgR基因的表达。他们将BmVgR3’UTR 构建至psiCHECKTM-2 海肾荧光素酶报告基因载体中,然后用这些miRNA 类似物(mimic)处理发现其中两个miRNAs 分别使荧光素酶活性降低了33.4%和38.4%,当两种miRNAs 同时存在时抑制作用更强,说明它们可能通过结合BmVgR3’UTR 上的作用位点来调控BmVgR基因的表达[28]。
4 启动子活性研究系统的发展与展望
目前而言,启动子活性的研究还非常依赖报告基因系统技术的发展,而荧光素酶报告基因系统较其他报告基因系统具有更多的优点和可操作性。同时也得益于生物技术和科学的飞快发展,萤火虫荧光素酶报告基因系统将会拥有更多的提升和进步空间。荧光素酶报告基因在应用过程中有以下几点优势:(1)在使用过程中不涉及放射性,使用安全污染;(2)荧光素酶检测信号强,比CAT(cat reporter assay)等其他报告基因检测速度更快,灵敏度高将近100~1000 倍[32];(3)样品检测步骤简单,操作方便;(4)比显微镜检测荧光更灵敏,没有非特异性激发光干扰,信噪比高不易受底物影响;(5)荧光素酶的半衰期短[33];(6)检测浓度线性范围广,只要荧光素酶的浓度在10-16mol/L(10pg/L)到10-8mol/L(1mg/L)内均合适检测[32]。
但双荧光素酶报告基因在应用过程中也有诸多注意事项,例如:(1)为了良好的测定效果,在双荧光素酶测试试剂与测试样品混合到测定的这段时间要保持一致,避免过长地停滞(要在1~2 s 时间内进行测定);(2)Renilla 荧光素酶检测液不能长期保留,配制后要立即使用;(3)荧光素底物反应液配制后不能反复冻融,不同批次的配制液可能存在差异;(4)温度对酶有很大影响,要保证样品和试剂测定时完全达到室温[17,20]。不过,当前也有些单个载体(如pmirGLO)能够同时携带两个不同的荧光素酶报告基因,还有各种功能性载体能满足不同实验的需求。
随着人类对基因启动子活性研究的逐渐深入,对于实验工具和研究体系的要求也会进一步提高,启动子活性研究系统亦会得到进一步的发展。随着科技的进步,实验操作和应用技术也将愈加熟练可控,新型技术运用范围也会更加广泛。可以预见,人类的智慧的发展和科学技术的飞跃会使双荧光素酶报告基因系统变得更加便利、高效,从而获得更广、更深的发挥空间,直至该项技术被更好的启动子活性研究系统所取代。
