黄曲霉对花生和玉米的侵染研究
2022-05-13荆丹岳晓凤白艺珍丁小霞张奇李培武
荆丹,岳晓凤,3,4*,白艺珍,3,4,丁小霞,3*,张奇,3,4,李培武,3,4
(1.中国农业科学院油料作物研究所,湖北 武汉, 430062;2.农业农村部油料产品质量安全风险评估实验室(武汉),湖北 武汉, 430062;3.农业农村部生物毒素检测重点实验室,湖北 武汉, 430062;4.国家农业检测基准实验室(生物毒素),湖北 武汉, 430062)
黄曲霉(Aspergillus flavus)是一种常见的腐生真菌,属子囊菌门发菌科曲霉属,其最适生长温度范围为25~42℃,最适产毒温度为28℃,适宜水分活度(water activity,aw)为0.86~0.96[1,2]。黄曲霉在世界各地广泛分布,既是一种植物病原菌,又是条件性的人畜共患病菌[2,3]。黄曲霉寄主范围广,能侵染玉米、花生、棉籽、大豆、坚果等多种农作物,进而诱发花生曲霉病、玉米穗腐病、棉花棉铃曲霉病等病害[4],其中对花生和玉米危害最为严重。
黄曲霉侵染作物后产生的黄曲霉毒素(aflatoxin, AFT)、环匹阿尼酸(cyclopiazonic acid, CPA)、黄曲霉震颤素(aflatrem)等次级代谢产物,是威胁农产品及食品质量安全的重要危害因子。其中,黄曲霉毒素B1是毒性最大、致癌力最强的真菌毒素,1993年被世界卫生组织(World Health Organization,WHO)国际癌症研究机构(International Agency for Research on Cancer,IARC)划定为I类致癌物。黄曲霉毒素污染不仅对粮食作物造成巨大的经济损失,而且对消费者的健康产生不利影响。据联合国粮食及农业组织(Food and Agriculture Organization of the United Nations,FAO)报道,全球每年受霉菌及其毒素污染的农作物约达25%,其中约2%的农产品因毒素污染超标而失去利用价值[5]。2016年,Mitchell 等评估,黄曲霉毒素污染导致美国玉米行业每年的经济损失高达5210 万至16.8 亿美元[6]。我国肝癌流行病学调查研究发现,一些地区人群膳食中黄曲霉毒素的污染水平与原发性肝癌的发生率呈正相关[7]。2014 年,Li 等对我国黄曲霉毒素污染调查研究显示,在长江三角洲地区采集的粮油产品(水稻、小麦、花生、玉米、大豆、燕麦、食用油)中,有14.5%受到了黄曲霉毒素的污染[8]。2015 年,Ding等发现中国长江流域主产区的花生黄曲霉毒素污染风险相对较高[9]。目前,有超过99 个国家已经采取行动控制农产品和食品中黄曲霉毒素的产生[10]。
黄曲霉能产生无性分生孢子和菌核两类繁殖体,其中分生孢子是主要的初侵染源,菌核能在极端条件下存活,有助于黄曲霉度过不利的生存环境。研究表明黄曲霉的生长发育及其次生代谢物的产生与多种环境因素有关,温度和水分活度是最主要的环境因子,高温和干旱有利于黄曲霉菌侵染和毒素污染的发生[11]。随着全球气候变暖、干旱频发等气候问题的加重,因黄曲霉侵染农作物引起的黄曲霉毒素污染已成为危及农产品质量安全和人类健康的全球性问题,发展中国家尤为严重[12~14]。因此,明确黄曲霉定殖、侵染过程及致病分子机理,对黄曲霉毒素污染早期预警以及源头防治策略的研发具有重要的指导意义。本文主要对花生、玉米黄曲霉的侵染过程、定殖、影响因素和侵染机制等进行了详细的综述,旨在为农作物中黄曲霉毒素污染预警与防控提供依据。
1 黄曲霉侵染循环
黄曲霉主要分布在纬度26°~35°的温暖气候区域,而在纬度45°以上地区很少发现[15]。土壤是黄曲霉的主要越冬场所,在自然条件下,黄曲霉通常以其无性繁殖体分生孢子或休眠体菌核存在于土壤中,而在植物组织中以菌丝的形式存在。菌核能在恶劣的环境下生存于土壤中,以菌丝的形式萌发生长并产生分生孢子,在炎热和干燥的气候条件下大量繁殖[16,17]。黄曲霉在农作物中的生活史主要包括两个阶段:一是黄曲霉菌定殖于土壤中的植物残体,以分生孢子或以菌核形式在土壤中越冬,或以菌丝体形式在植物残体上越冬,二是侵染生长期的谷物或种子[18]。
黄曲霉的侵染开始于分生孢子在植物体上定殖。土壤中的生物运动及雨水的冲刷最易导致黄曲霉在花生种子中定殖、生长及繁殖[19,20],而空气传播的黄曲霉分生孢子对玉米、坚果等地上作物的侵染更为重要。在黄曲霉的整个侵染循环中,主要包括气生菌丝、菌丝体、分生孢子、菌核、子囊孢子等组织结构[21]。
花生由于其地下结实的特性,花生荚果与土壤中的黄曲霉直接接触。花生播种后,随着温度逐渐升高,土壤中的黄曲霉分生孢子或菌核在适宜条件下萌发长出新的菌丝、并产生大量分生孢子,通过不同方式进行侵染,如通过花、果针或系统侵染。但侵染主要发生在荚果周围的土壤中。当果针下扎到土壤后,菌丝或分生孢子通过荚果壁侵染发育中的果实和种子。当土壤翻动或受雨水冲刷时,土壤中分生孢子快速传播从而引发更加广泛的侵染[18,22]
与花生黄曲霉侵染不同,玉米黄曲霉侵染需要借风、雨水、气流、昆虫等媒介将土壤中黄曲霉分生孢子传播至地上作物后开始初侵染。黄曲霉的分生孢子可以定殖于玉米的穗轴和穗须。幼嫩的玉米穗轴在灌浆后期(早期干旱阶段)最容易感染黄曲霉的孢子,而成熟期的玉米穗须则更容易受侵染[23]。生长期间的植株受到黄曲霉侵染后,主要以菌丝体在植物组织内生长,到一定的生长阶段或在某些环境因素诱导的条件下,黄曲霉形成无性孢子或者菌核,再次回到土壤中,或者扩散到其他健康植株,开始下一轮的侵染循环[21],导致病害扩展流行。
2 花生黄曲霉侵染
花生是我国重要的油料作物和经济作物,产量居我国油料作物首位,占世界花生总产的40%。与大豆等其他油料作物相比,花生是最易受黄曲霉侵染及黄曲霉毒素污染的油料作物[24]。花生作为地上开花地下结果的作物,其果实可与土壤中的黄曲霉直接接触而受到侵染[25]。研究表明,土壤是黄曲霉的主要栖息地,也是花生黄曲霉的主要来源[26,27],花生荚果中的黄曲霉与土壤中的黄曲霉有直接联系[20]。
2.1 花生黄曲霉定殖及侵染过程
黄曲霉侵染和黄曲霉毒素的产生是真菌、宿主相互作用的结果。黄曲霉穿透花生果壳、种皮,最终在花生子叶中定殖。花生子叶含有丰富的油脂和蛋白质,可以为黄曲霉的生长、繁殖与代谢提供充足的营养物质。Nayak[28]等利用荧光显微镜对黄曲霉离体接种的抗病和感病花生种子不同时间点的侵染情况进行研究,发现接种1 d 时,抗病花生种子上的菌丝生长相对较慢;2 d 时,菌丝在感病花生种子上的生长速度明显高于抗病花生种子;3 d 时,这种趋势更为明显;7 d 时,在感病花生种子上能观察到明显的产孢,而抗病花生种子上只有很少的分生孢子。表明无论是感病还是抗病花生种子在接种黄曲霉后,第3 d 均可清楚地观察到菌丝,第7 d孢子在种皮表面形成。闫彩霞等[29]研究发现,黄曲霉侵染抗病花生3 d、6 d 和9 d 后,花生胚细胞壁和细胞膜依然比较完整,脂肪粒成团聚集存在,而感病花生的胚细胞壁和细胞膜则逐渐裂解,脂肪粒大量流失,侵染9 d 后,部分细胞的脂肪粒已完全消失。表明在花生胚表皮出现肉眼可见的黄曲霉之前,其籽粒内部的细胞结构和化学成分已经发生质变。Sobolev 等[30]研究了花生被黄曲霉侵染后,通过从被侵染区域到健康组织对花生不同层间和不同深度进行解剖。结果发现在花生籽仁切片中检测到6种花生抗毒素,即反式白藜芦醇、反式花生四烯醇-1、反式花生四烯醇-2、反式花生四烯醇-3、反式3-异戊二烯- 4,3,5 -三羟二苯乙烯和SB -1。黄曲霉的侵染可以导致植物抗毒素的产生,但植物抗毒素的种类在不同层间有显著差异,在侵染24 h 后,远离感染区域的组织几乎只产生反式白藜芦醇,而靠近感染区域的组织则合成全部6 种植物抗毒素;48 h 后,在花生更深层组织处检测到了6 种植物抗毒素。该研究初步阐明了在黄曲霉侵染花生籽粒过程中植物抗毒素合成的动力学规律,为进一步探究黄曲霉侵染花生籽粒的机理及培育抗黄曲霉花生品种提供了一定的理论依据。最新研究发现,哌啶酸是花生抵御黄曲霉侵染的重要因子,外源添加哌啶酸显著降低黄曲霉对感病花生的侵染率[31]。
2.2 花生黄曲霉侵染影响因素
黄曲霉可以通过不同的途径侵染处于生长发育时期的花生,但是侵染主要发生在荚果与土壤直接接触的部位。影响因素主要有干旱和高温胁迫、土壤质地、病虫害等。
灌浆期间的天气条件对作物产量和品质均有显著影响,特别是黄曲霉毒素的污染。在充足的降雨或灌溉条件下,黄曲霉毒素的产生几率较小,然而世界上主要的花生种植区生长条件并不理想[32]。Sanders 等[33]研究表明当花生荚果暴露在干旱条件下,即使根部水分供应充足,花生也会受到黄曲霉的污染。Craufurd 等[34]证实当土壤温度接近黄曲霉的最佳生长条件时,黄曲霉对花生的侵染和花生中黄曲霉毒素的含量与灌浆期土壤中发生水分胁迫有关。一方面,干旱胁迫降低了花生对黄曲霉侵染的天然防御能力[35]。另一方面,土壤中水分活度的降低会抑制细菌、变形虫以及竞争性真菌的生长和繁殖,从而加快了适宜该条件的黄曲霉菌的生长[32]。如果花生在收获后没有及时干燥并保持在适宜的水分范围内,会进一步导致收获后黄曲霉毒素的污染和累积。此外,有研究表明不同土壤类型对花生的霉菌感染水平具有显著的影响。例如,沙质土壤含水量低,更有利于真菌的快速繁殖;而黏土有较高的持水能力,因此发生干旱胁迫的可能性较小,受到霉菌感染机率低[36]。害虫对花生荚果造成的伤口也为黄曲霉菌侵染创造了有利的位点,从而增加了黄曲霉侵染的几率和黄曲霉毒素污染发生的程度[37]。
3 玉米黄曲霉侵染
玉米是世界种植面积最广且最为重要的经济作物之一,亦是我国主要的粮饲兼用作物。玉米极易遭受黄曲霉侵染和黄曲霉毒素污染,不仅威胁人畜生命健康,还造成巨大的经济损失。受到黄曲霉毒素污染的玉米籽粒,经过加工或深加工残留在相关的制品中,如玉米食品、玉米油、玉米饲料等,给人体和动物健康造成危害[38,39]。Mitchell等研究发现玉米黄曲霉毒素污染导致美国每年大约7亿美元的经济损失[38]。
3.1 玉米黄曲霉定殖及侵染过程
在田间,黄曲霉侵染玉米最常见的侵染途径是通过空气传播。在玉米上的定殖起始于玉米穗须,然后产生分生孢子迁移到玉米穗部,在适宜的条件下萌发出菌丝,进而菌丝通过不断地延伸扩展到玉米籽粒,这个侵染过程大概需要8天[40,41]。待黄曲霉菌丝覆盖玉米籽粒后,进入无性繁殖阶段,最终在分生孢子梗上产生大量成串的分生孢子,进而传播到周围的玉米植株,造成大面积的黄曲霉病害。Dolezal 等[42]研究发现接种玉米籽粒24 h 后,胚乳组织中出现了黄曲霉;72 h后,黄曲霉出现在玉米胚芽中;在72~96 h 之间,玉米花梗组织开始被黄曲霉感染。通过显微观察,发现菌丝主要定殖在胚乳的接种点及其周围,进而向下生长至胚芽,然后菌丝在胚乳—胚芽界面分化并形成包围胚芽的生物膜结构。Shu 等[43]采用组织切片方法观察到黄曲霉接种玉米籽粒后,在4 h、12 h 和24 h 时没有在组织中发现黄曲霉;而48 h 时,黄曲霉在玉米籽粒的糊粉层和胚乳最外层广泛定殖;糊粉层的破坏与黄曲霉菌丝的存在有关,72 h时,黄曲霉菌丝在玉米胚乳与胚芽交界处出现。
3.2 玉米黄曲霉侵染影响因素
黄曲霉侵染玉米除了田间玉米抽穗期间,由于害虫危害如玉米钻孔虫等危害产生伤口、土壤贫瘠及土壤干旱引发的穗部侵染[44],还会以为种子收获后储存条件不当造成侵染。玉米产量与黄曲霉侵染的关联性受一系列环境因素的影响,包括昆虫量、天气(降雨量、温度和湿度)及土壤类型[45~48]。在玉米抽穗期受到鸟类、昆虫、机械损伤等条件影响下,会导致黄曲霉侵染加重[49~51]。如玉米穗被昆虫咬食会造成组织的机械损伤,从而使黄曲霉孢子定殖在玉米伤口上,增加黄曲霉侵染的机率[52~54]。Yu等[55]研究表明,在美国转基因Bt 玉米能减少玉米螟的危害,玉米受虫口伤害减少,从而导致黄曲霉侵染及黄曲霉毒素污染率降低。另外,气候条件也是影响黄曲霉侵染水平的主要因素。黄曲霉毒素在玉米中的积累往往与气候因子对植株的胁迫有关。例如,干旱和高温是增加黄曲霉侵染风险和降低玉米产量最常见的气候因素[46,47,56,57]。干旱胁迫与玉米虫害发生的增加、黄曲霉侵染以及黄曲霉毒素积累有关[54,58]。干旱会引起玉米的防御能力减弱,发生虫害的风险会随之增加。高温可以加速玉米水分流失[52],从而可能导致氮和钾缺乏,玉米无法调动营养物质进行及时补给,对外界的防御能力下降[59]。作物收获后,贮藏环境(温度、湿度、贮藏害虫等)和贮藏时间是影响玉米收获后黄曲霉毒素污染的主要因素。入库贮藏的玉米所携带黄曲霉菌量的基数越大,就越易诱发黄曲霉毒素的污染。而破损、病残的玉米籽粒是黄曲霉最有可能的携带者,库存玉米籽粒回潮(种子含水量≥10%)或是仓库温度过高(≥20℃)会加快黄曲霉的生长和繁殖,从而加大黄曲霉毒素污染发生的机率。另外,玉米籽粒自身的代谢能力随着贮藏时间的增长而衰弱,抵抗黄曲霉侵染的能力下降,发生黄曲霉毒素污染的几率增加[60]。
4 黄曲霉侵染分子机制研究
在黄曲霉生长发育和侵染农作物过程中,涉及各类基因表达的调控。2006 年,Payne 等公布了黄曲霉全基因组序列信息,基因组大小约37 Mb,由8条染色体组成,预测含有12 000 多个功能基因[61,62],为黄曲霉功能基因组学的研究提供了遗传信息。随着分子生物学技术的快速发展和真菌遗传转化体系的成熟,黄曲霉侵染及致病相关基因研究已经取得初步进展,已有一些黄曲霉侵染、致病相关基因得到鉴定和功能分析,进一步深化了对黄曲霉侵染和致病分子机理的认识。
目前,黄曲霉侵染及致病相关基因研究已经取得初步进展,植物病原真菌的异三聚体G 蛋白在营养生长、产孢和致病等许多生物学过程中起着关键的调控作用。Liu 等[63]研究鉴定了黄曲霉中4 个G蛋白α 亚基编码基因(GanA、GpaB、FadA和GaoC),发现GpaB的缺失导致分生孢子产量下降,产孢相关基因brlA和abaA的表达显著降低。GpaB缺失突变体对花生种子的侵染能力明显减弱,且突变体细胞内cAMP 水平显著降低,表明GpaB作用于cAMP 信号途径,且在黄曲霉孢子形成、侵染等方面起着重要作用。pbsB基因是编码MAP(促分裂原活化蛋白)激酶级联反应中MAP 激酶的一种MAPKK(促分裂原活化蛋白激酶激酶),研究发现[64]pbsB缺失突变体的菌丝生长、分生孢子和菌核形成与野生型菌株相比受到明显的抑制,且对花生籽粒的侵染能力显著下降,表明MAPK(促分裂原活化蛋白激酶)信号途径在黄曲霉的营养生长、产孢、菌核形成、侵染和致病等过程中起着重要的作用。Zhang 等[65]研究发现MAPK 编码基因AflSlt2在黄曲霉、生长、产孢、致病以及毒素合成过程中具有重要的作用。AflSlt2缺失突变体完全丧失对花生的侵染力。磷酸酶介导的去磷酸化是细胞信号通路的组成部分,在真核生物各种细胞调控过程中发挥着重要作用,Yang 等[66]研究发现黄曲霉双特异性磷酸酶基因AflCDC14的缺失导致黄曲霉营养生长减慢,分生孢子形态异常,且不能产生菌核,表明AflCDC14磷酸酶可能在黄曲霉营养生长、菌核形成和致病等方面起着重要作用,并可作为黄曲霉防治的潜在药剂靶点。组蛋白乙酰转移酶(Histone Acetyltransferases,HATs)对真菌的生长发育和次生代谢产物的产生起着重要的调节作用,Lan 等[67]发现黄曲霉组蛋白乙酰转移酶编码基因AflGcnE参与调控黄曲霉的营养生长、产孢、菌核形成和花生种子定殖等过程。分生孢子色素是真菌细胞壁的重要成分,对真菌的生长发育极为重要,魏鹏霖等[68]鉴定了黄曲霉分生孢子色素合成基因pks1,发现pks1基因的缺失突变体对花生种子的侵染能力下降,明确了pks1基因是黄曲霉分生孢子色素合成的关键基因,影响黄曲霉的营养生长、产孢、菌核形成和对花生种子的侵染能力。Asis等[69]通过采用诱导培养基诱导黄曲霉产生胞外蛋白酶的方法,明确了黄曲霉产生的金属蛋白酶和丝氨酸蛋白酶参与花生种子的侵染,导致种子组织损伤,种子活力下降,进而促进黄曲霉通过种皮侵入花生种子。多胺(PAs)是影响作物生长发育和应激反应的多聚体,在真菌致病过程中起着重要作用,其中亚精胺(Spd)的生物合成在黄曲霉的生长发育和致病过程中起着关键性作用。Majumdar等[70]研究发现,在黄曲霉侵染玉米籽粒过程中,黄曲霉胞内亚精胺合酶基因的缺失对黄曲霉生长、多胺合成基因的表达产生了不利的影响。与野生型相比,Spd缺失突变体导致黄曲霉的生长、产孢和黄曲霉毒素产量显著下降,表明Spd 的生物合成对黄曲霉的正常发育和致病机制至关重要。Yang 等[71]采用基因替换策略对黄曲霉腺苷酸环化酶编码基因acyA进行了敲除,结果发现黄曲霉acyA缺失突变体的营养生长、产孢和菌核形成等方面存在严重缺陷,表明acyA基因在黄曲霉的营养生长、产孢、致病和毒素生物合成过程中起着重要作用。Yuan 等[72]研究发现hexA基因的缺失导致黄曲霉在玉米籽粒上的产孢量及其产毒力显著降低,表明hexA基因在黄曲霉分生孢子的产生、侵染、致病和毒素合成等过程中起着重要作用。Tumukunde 等[73]研究发现,AfsakA突变菌株能产生更多的分生孢子和黄曲霉毒素B1,表明AfsakA基因存在可以导致黄曲霉的孢子和毒素产量下降。这些研究均为黄曲霉新的防治手段的研发提供理论支持,也为新药剂靶标的寻找奠定理论基础。
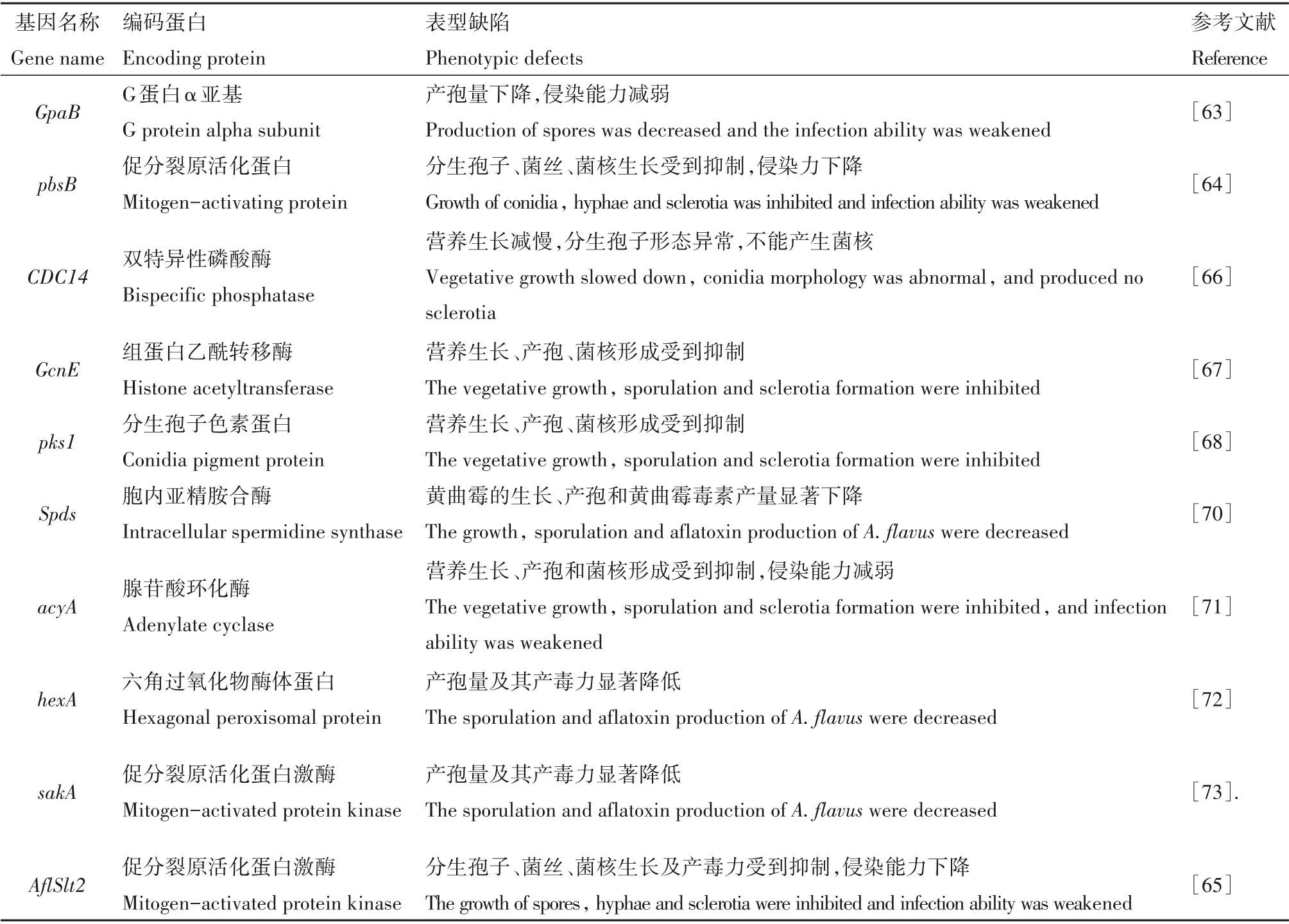
表1 黄曲霉侵染及致病相关基因Table 1 Infection and pathogenicity-related genes of A.flavas
5 展望
花生、玉米等粮油作物在田间生产、收储、运输等过程都有可能受到黄曲霉菌侵染和黄曲霉毒素污染。黄曲霉侵染和毒素发生是病原菌、寄主植物和环境三者相互作用、相互影响的结果。目前,选育抗黄曲霉侵染、抗黄曲霉产毒的抗病品种和田间绿色生物防治等方法是黄曲霉污染防控的有效手段。近年来研究发现,花生荚壳抵御黄曲霉侵染的能力强弱对降低毒素污染具有重要作用,发掘荚壳与种子侵染抗性、种子产毒抗性整合的抗性优质材料是培育突破性的抗黄曲霉品种的重要途径[74]。然而,相同或不同地理来源的黄曲霉菌株间侵染力与产毒力均存在显著差异[75,76],增加了抗性品种选育和毒素污染控制难度。因此,进一步以花生-黄曲霉或玉米-黄曲霉为研究对象,结合基因组、代谢组等多组学整合分析技术,研究不同侵染力与产毒力菌株间遗传变异机制,深入发掘黄曲霉侵染和产毒调控基因,探明调控网络,揭示黄曲霉的侵染、致病和产毒机理及其与寄主、环境的互作关系,为花生、玉米抗黄曲霉侵染和产毒品种材料选育,新型防治靶标筛选以及源头防控策略研发奠定理论基础,同时也为其他农作物中黄曲霉侵染及黄曲霉毒素污染防控提供参考借鉴。
