大肠杆菌噬菌体抗性菌株与其敏感菌株的比较研究
2022-04-25崔嘉琪孙蓉蓉刘文华任慧英
崔嘉琪,孙蓉蓉,张 灿,刘文华,邹 玲,任慧英
(青岛农业大学动物医学院,山东 青岛 266109)
早在上世纪30~40 年代噬菌体就被用于治疗细菌感染,但随着抗生素的发现,因其高效杀菌、使用方便、价格便宜的特点,抗生素很快在世界各地得到广泛应用,逐渐代替了噬菌体。近年来,由于抗生素的滥用致使细菌耐药性产生甚至不断出现了多重耐药菌,寻找抗生素的替代品是目前的当务之急。噬菌体是能够特异性感染细菌的病毒[1],具有特异性的宿主识别谱,可实现对细菌的快速特异杀灭,其杀菌作用不受细菌耐药性的限制,且噬菌体具有研发周期短、投入成本低、安全性高等优点,成为最有影响力的抗生素替代品之一[2-3]。因此噬菌体疗法的研究及临床应用逐渐得到重视。
在细菌与噬菌体共同进化过程中,细菌不断演化出抵抗噬菌体的机制,噬菌体抗性菌的出现成为噬菌体治疗中的难题[4],噬菌体的抗性菌会不会像抗生素抗性菌一样最终面临无药可治的境地?噬菌体抗性细菌的出现往往与细菌的自发突变和适应性有关,且往往与噬菌体吸附受体的修饰有关。主要的噬菌体抗性机制是噬菌体抗性菌株能够通过表面修饰对噬菌体进行防御[5]。乐率研究发现,绿脓杆菌通过细胞表面外膜脂多糖的丢失从而产生了噬菌体抗性,且抗性菌的毒力显著下降[6]。本实验室前期分离到一株大肠杆菌噬菌体Bp4,属于短尾噬菌体科N4 类噬菌体,能够裂解O78 血清型的大肠杆菌。本研究通过双层平板法筛选大肠杆菌噬菌体的抗性突变株,将其与大肠杆菌敏感菌株进行了形态及培养特性观察、血清型及耐药性检测、基因组测序分析等研究,以期为噬菌体治疗中噬菌体抗性菌的研究提供参考依据。
1 材料与方法
1.1 主要实验材料大肠杆菌噬菌体Bp4、大肠杆菌血清O78 型敏感菌株(后简称O78 型敏感菌株)均由本实验室保存;BALB/c 小鼠购自青岛大任富城畜牧有限公司。营养琼脂、LB 肉汤、琼脂粉均购自生工生物工程(上海)有限公司;大肠杆菌O 抗血清购自中国兽医药品监察所;药敏纸片购自杭州滨和微生物试剂有限公司。
1.2 噬菌体抗性突变菌株的筛选与鉴定将噬菌体Bp4 的宿主菌(O78 型敏感菌株)接种于5 mL LB 液体培养基中,37 ℃220 r/min 振荡培养至对数生长期后取200 μL 与200 μL 噬菌体Bp4 的增殖液一起加入5 mL LB 中,37 ℃220 r/min 振荡培养,观察液体的浑浊状态;在约3 h 液体清亮后继续振荡培养过夜至浑浊,用接种环采用三线法将该噬菌体增殖液接种于LB 平板,37 ℃倒置培养过夜。将LB 平板上长出的单菌落连续纯化4 代,将第4 代纯培养的单菌落再与噬菌体Bp4 经液体培养后再经双层平板法培养[7],即用接种环挑取第4 代纯培养的单菌落接种于5 mL LB 中,37 ℃220 r/min 振荡培养至对数生长期,取110 μL 菌液与110 μL 噬菌体Bp4 增殖液混匀后于37 ℃孵育5 min,取200 μL 孵育液加至已溶解的上层琼脂中,混匀后迅速倾倒在下层营养琼脂上,晃匀待凝固后37 ℃倒置培养6 h~8 h,设不加噬菌体的敏感菌株为阳性对照。观察噬菌斑产生情况,不产生噬菌斑的菌株即为对噬菌体Bp4 具有抗性的大肠杆菌突变菌株,命名为大肠杆菌O78-R 型抗性菌株(后简称O78-R 抗性菌株)。
1.3 敏感菌株与抗性菌株的形态及培养特性观察将O78 型敏感菌株与O78-R 抗性菌株分别在普通营养琼脂及麦康凯琼脂上划线培养,37 ℃16 h 后观察菌落形态,挑取单菌落经革兰氏染色观察。挑取单菌落于5 mL LB 液体培养基中,37 ℃220 r/min 振荡过夜培养,利用倾注平板法[8]测定菌液浓度:将菌液10 倍倍比稀释至107倍,分别选取105和107稀释度的菌悬液各1 mL 于无菌培养皿中,然后将约50 ℃营养琼脂注入平板中晃匀,试验重复两次;待琼脂凝固后,将平板倒置于37 ℃温箱中培养24 h 左右,经细菌计数评估两种细菌的生长速度。
1.4 敏感菌株与抗性菌株的基因组测序及序列分析将纯化的O78 型敏感菌株及O78-R 抗性菌株由北京安诺优达基因科技有限公司测序,根据测序结果分析比较抗性菌株与敏感菌株基因组的差异。
1.5 敏感菌株与抗性菌株的血清型鉴定将O78 型敏感菌株与O78-R 抗性菌株分别接种于5 mL LB 肉汤中,37 ℃220 r/min 振荡培养12 h 后,4 000 r/min离心10 min,弃上清,用0.5%石炭酸生理盐水2 mL悬浮后121 ℃高压2 h,以破坏细菌的荚膜“K”抗原,制成高压抗原,以大肠杆菌O 抗血清为抗体,利用玻板凝集法鉴定大肠杆菌的“O”抗原血清型。
1.6 敏感菌株与抗性菌株的耐药性检测参照文献[9]采用药敏K-B 纸片法分别检测O78 型敏感菌株及O78-R 抗性菌株对12 种药物的敏感性,根据欧盟药敏试验标准判定结果。
1.7 敏感菌株与抗性菌株的致病性试验将20 只BALB/c 小鼠随机分为两组,每组10 只,分别腹腔注射O78 型敏感菌株新鲜菌液及O78-R 抗性菌株新鲜菌液0.2 mL(菌液浓度均为1×108cfu/mL),统计感染后72 h 内两组小鼠的发病率和死亡率,并统计感染后12 h、24 h、36 h、48 h、72 h 时每组小鼠的临床症状得分,判定标准为:死亡0 分,濒临死亡1分,部分闭眼、眼周有渗出液2 分,嗜睡、弓背3分,运动减少、被毛粗乱为4 分,正常、健康者5分,绘制患病小鼠的计分及其存活率曲线。
2 结 果
2.1 噬菌体抗性菌株的筛选与鉴定结果分别取200 μL 培养至对数期的O78 型敏感菌株菌液及噬菌体Bp4 增殖液于5 mL 肉汤中,振荡培养,可观察到液体从清亮-浑浊-清亮-浑浊的变化,取最后浑浊的液体划线传代,用噬菌体Bp4 再感染第4 代的菌液,液体一直处于浑浊状态,初步判断噬菌体Bp4不再感染该菌株;用第4 代的菌液与噬菌体Bp4 经双层平板培养6 h~8 h 不再产生噬菌斑,而O78 型敏感菌株的平板则产生了明显的噬菌斑(图1)。表明宿主菌(敏感菌株O78 型)在与噬菌体Bp4 相互作用中产生了对噬菌体Bp4 耐受的菌株,即抗性菌株O78-R。
图1 噬菌体Bp4抗性菌株的筛选与鉴定结果Fig.1 Screening and identification of E.coli strains resistant to phage Bp4
2.2 敏感菌株与抗性菌株的形态及培养特性结果将抗性菌株与敏感菌株进行了形态及培养特性的比较,发现两株菌的菌落形态相同,在普通营养琼脂培养基上均呈表面光滑、半透明的单菌落(图2),在麦康凯培养基上呈粉红色小菌落,表面光滑。且抗性菌株与敏感菌株的生长速度相同,105稀释度的两株菌菌悬液生长的单菌落数量太多不予计数,107稀释度两株菌的菌悬液在37 ℃培养24 h 时均生长出30 个单菌落,且重复实验结果相同。表明,抗性菌株与敏感菌株的形态及培养特性相同,生长速度一致。
图2 O78敏感菌株与O78-R抗性菌株在普通营养琼脂培养基的培养结果Fig.2 Culture results of O78 sensitive strain and O78-R resistant strain in common nutrient agar medium
2.3 敏感菌株与抗性菌株基因组差异的分析将敏感菌株与抗性菌株分别测序。结果显示,O78 型敏感菌株与O78-R 抗性菌株基因组全长均为4 798 435 bp。两组测序数据的比对分析结果显示,抗性菌株中有3 个碱基发生突变,分别位于3 个重叠群(contig)中(表1)。
表1 显示,3 个碱基的突变涉及到3 个氨基酸,分别为乙醛脱氢酶的N→Y,十一碳二烯磷酸α-N-乙酰氨基葡萄糖磷酸转移酶1 的W 发生无义突变,以及UpfB 蛋白L 发生同义突变。致噬菌体发生抗性的原因很可能为contig00024 中65 018 nt 处的碱基突变,密码子由TGG 变为TAG,编码的氨基酸由色氨酸突变为终止密码子,从而致蛋白质翻译提前终止,致使抗性菌株的葡萄糖磷酸转移酶由敏感菌株的372 个氨基酸缩短为133 个氨基酸,不能执行正常的酶功能(图3)。

表1 抗性菌株突变点的信息Table 1 Annotation for mutantion sites of the resistant strain

图3 敏感菌株O78与抗性菌株O78-R氨基酸序列比对Fig. 3 Comparison of amino acid sequence between the sensitive strain O78 and resistant strain O78-R
2.4 敏感菌株与抗性菌株的血清型鉴定采用玻板凝集法对O78 型敏感菌株及O78-R 抗性菌株进行血清型鉴定,结果显示,O78 型敏感菌株与O78 抗血清呈明显的凝集现象,液滴边缘出现麸皮样颗粒状的凝集片,液体清亮;O78-R 抗性菌株与O78 抗血清的凝集现象不明显,液体浑浊,且有少量细微颗粒。表明敏感菌株确为O78 血清型,而抗性菌株血清型发生了改变,已不能鉴定为O78 血清型。
2.5 敏感菌株与抗性菌株的药敏试验采用药敏纸片法分别检测O78 型敏感菌株与O78-R 抗性菌株的药物敏感性。结果显示,与敏感菌株相比,O78-R抗性菌株对替米考星、环丙沙星、氨曲南、妥布霉素的抑菌圈直径增大,耐药性降低(表2),表明噬菌体抗性菌株比其敏感菌株对以上抗生素更敏感。
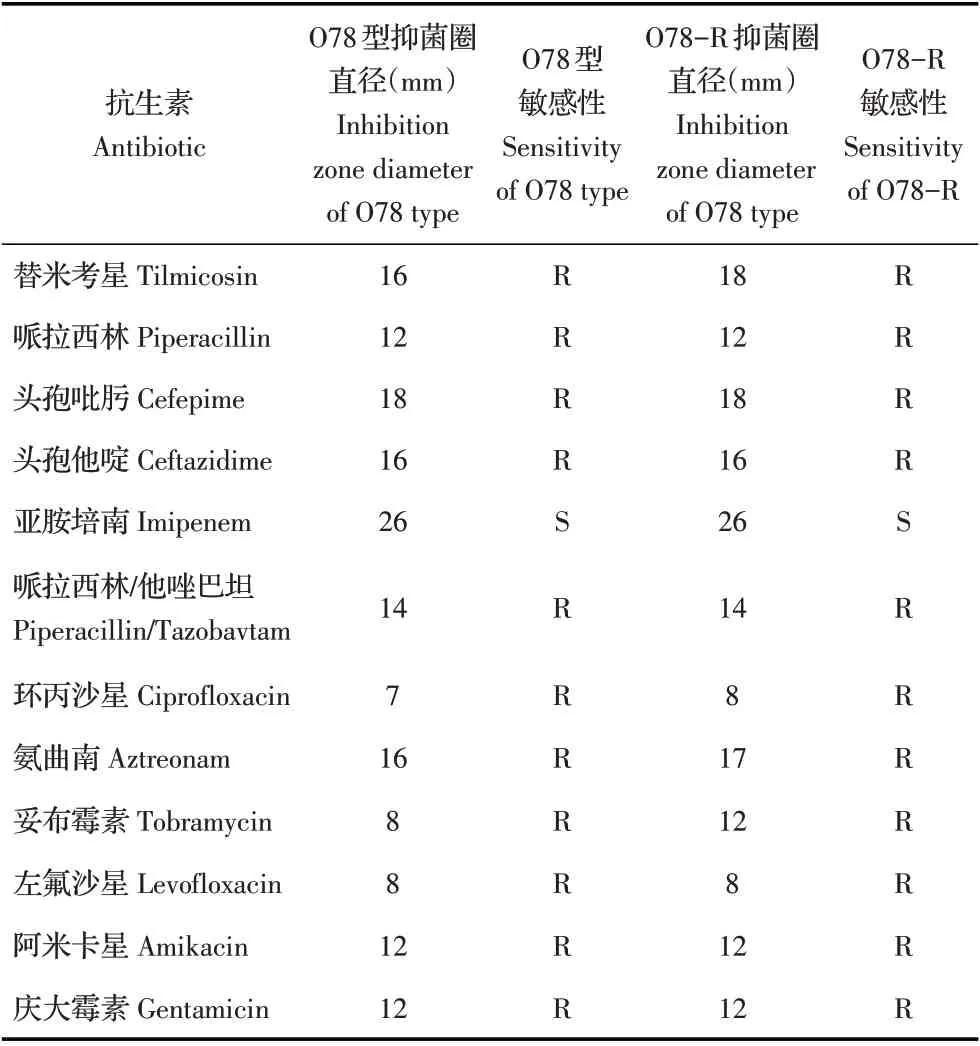
表2 药敏试验结果Table 2 Results of drug sensitivity test
2.6 敏感菌株与抗性菌株对小鼠的致病性试验结果将敏感菌株与抗性菌株的增殖液经腹腔接种小鼠,记录小鼠的体况计分及存活率。结果显示,抗性菌株接种的小鼠未出现死亡,在感染后0~48 h 小鼠体况计分一直维持在45 分以上,之后略有下降(图4A);而敏感菌株接种的小鼠在接种后72 h 内共死亡4只,体况计分比抗性菌株接种的小鼠约低30分(图4B),表明噬菌体抗性菌株对小鼠的致病性降低。
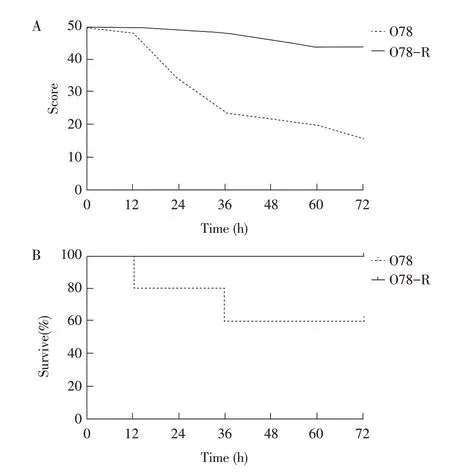
图4 两组小鼠的体况计分(A)及存活率(B)统计结果Fig.4 Statistical results of body condition score(A)and survival rate(B)of two groups of mice
3 讨 论
为了研究噬菌体的抗性突变大肠杆菌菌株与其敏感菌株的差异,本研究经双层平板法筛选出对噬菌体具有抗性的大肠杆菌突变菌株,虽然在形态及培养特性上该抗性菌株与其敏感菌株未表现出差异,但该抗性菌株不再与O78 抗血清发生强阳性的凝集反应,不再属于O78 血清型。
基因组测序及分析结果显示,敏感菌株与抗性菌株基因组的全长一致,证明抗性菌株并无基因缺失和插入,但在抗性菌株中出现3 处碱基的突变,导致1 个同义突变,1 个氨基酸替换(乙醛脱氢酶)以及1 个无义突变。乙醛脱氢酶负责催化乙醛氧化为乙酸的反应,参与细菌的代谢,与噬菌体的感染无关。无义突变导致抗性菌株的十一碳二烯磷酸α-N-乙酰氨基葡萄糖磷酸转移酶1 的翻译提前终止,而该蛋白参与细菌外膜中脂多糖O 抗原的生物合成途径[10],导致脂多糖O 抗原的合成受到抑制,提示该蛋白的突变是导致噬菌体不能感染抗性菌株的原因。本研究所用的噬菌体Bp4 属于短尾噬菌体科,为N4 类噬菌体[11],短尾噬菌体中N4 类噬菌体G7C识别大肠杆菌表面的脂多糖受体[12],T7 类噬菌体尾部蛋白识别的也是宿主菌表面的脂多糖受体[13],推测脂多糖也是噬菌体Bp4 的受体。大肠杆菌为革兰氏阴性菌,细胞壁外膜中镶嵌大量的脂多糖[14]。革兰氏阴性菌的脂多糖由3 部分组成:从内向外依次为脂质A、核心多糖、多糖侧链,多糖侧链又称为O-抗原,当脂多糖中的任何一部分缺失均会对脂多糖的合成造成影响。本实验中筛选的抗性菌株是由于碱基突变导致了色氨酸变为终止密码子,致脂多糖中参与O 抗原生物合成的N-乙酰葡萄糖胺磷酸转移酶的翻译提前终止。通过凝集试验鉴定敏感菌株与抗性菌株的O 抗原,结果显示敏感菌株为典型的O78 血清型,而抗性菌株不再与O78 抗血清发生凝集,不能鉴定为O78 血清型。该结果证实O 抗原是噬菌体Bp4 识别并吸附宿主菌的关键受体。大肠杆菌的O 抗原类型与其致病性有直接相关性,每株大肠杆菌只含有一种O 抗原,能致鸡大肠杆菌病的常见血清型为O1、O2、O35 及O78[15]。本研究获得的噬菌体抗性菌不再具有O78 抗血清的凝集性,提示细菌的致病性可能下降。乐率等通过对铜绿假单胞菌PA1 株与噬菌体的相互作用,筛选出抗性菌株PA1r 并做了相关研究,发现抗性菌株可以分泌红色素,且致病性下降[6]。噬菌体抗性菌株致病性的下降已被多项研究所证实[4,16],本研究经腹腔注射敏感菌株小鼠的死亡率达到40%,而腹腔注射抗性菌株小鼠的死亡率为0,也证明了噬菌体抗性菌株致病性的下降。噬菌体抗性菌株致病性的下降会导致细菌在动物体内更容易被免疫系统清除。
本研究发现,噬菌体抗性菌株的耐药性发生了改变,其对替米考星、环丙沙星、氨曲南、妥布霉素的抑菌圈直径增大,比噬菌体敏感菌株的耐药性降低,该结果与Jalasvuori 报道的结果类似[17]。噬菌体抗性菌株耐药性的下降会导致其在较低的抗生素浓度下即可被杀死。
噬菌体抗性菌株产生的几率大约为千万分之一。细菌对噬菌体的抗性是相对的,其对一株噬菌体产生抗性,但还可以被其他的噬菌体感染。于玲等分离到对大肠杆菌具有抗性的噬菌体,并将其与原噬菌体混合制成新的噬菌体鸡尾酒用于杀菌,结果表明,该噬菌体鸡尾酒不仅具有明显的杀菌效果,而且在体外降低了噬菌体抗性菌株的产生和突变频率[18]。
噬菌体抗性突变菌株的产生是细菌和噬菌体相互作用的结果,抗性菌株的产生必然会导致细菌基因组及表型的变化。本研究通过分析大肠杆菌短尾噬菌体Bp4 的抗性菌株和敏感菌株,发现抗性菌株的耐药性和致病性均下降,说明噬菌体抗性菌株的产生不会对噬菌体治疗效果产生较多副作用,也为噬菌体治疗中出现抗性菌问题的解决提供了参考依据。
