iucA基因缺失对尿路致病性大肠埃希菌增殖、黏附、侵袭和定植能力的影响
2022-03-05康嘉乐林梅双林艳芬
邓 聪,康嘉乐,林梅双,林艳芬
尿路感染(urinary tract infections,UTI)是临床上最常见的细菌感染之一[1],其高居不下的复发率、再感染率以及日益增高的多重耐药菌的检出率,强调了发展UTI新的治疗策略的必要性[2]。约75%~95%的UTI由尿路致病性大肠埃希菌(UropathogenicE.coli,UPEC)所致[3]。铁摄取系统是UPEC目前已知的毒力因子之一[4-5]。
铁是微生物生命活动的必要元素。病原菌通过分泌铁载体与宿主体内的铁离子结合,形成复合物并通过外膜表面的相应受体摄取。UPEC可以分泌多种铁载体,包括耶尔森杆菌素、气杆菌素、肠杆菌素和沙门菌素[6]。其中气杆菌素已被证实通过促进菌株生长、定植等,在肺炎克雷伯菌、禽致病性大肠埃希菌等的致病过程中发挥重要作用[7-8],但其与UPEC致病之间的关系至今仍不明确。该研究通过对气杆菌素编码基因iucA缺失株和回补株的构建,分析iucA基因在UPEC致病中的作用。
1 材料与方法
1.1 实验材料
1.1.1菌株、质粒和细胞 大肠埃希菌CFT073菌株(临床UTI分离的大肠埃希菌,由美国南加州大学洛杉矶儿童医院黄胜和教授惠赠)。pKD46质粒、pKD3质粒、pCP20质粒、pRK415质粒和人膀胱癌上皮细胞株5637由本实验室保存。
1.1.2实验动物 24只C57BL/6雌性小鼠购自湖南斯莱克景达实验动物有限公司。等级:SPF级。体质量:18~24 g。随机分为3组(CFT073组、ΔiucA组、C-iucA组),每组8只。
1.1.3试剂与仪器 TaqPlus DNA聚合酶、质粒抽提试剂盒和胶回收试剂盒(大连宝生物工程有限公司);L-阿拉伯糖、氯霉素、氨苄青霉素、四环素和庆大霉素(美国Sigma公司)。 PCR扩增仪、电击仪(美国Bio-Rad公司)、分光光度计(美国Beckman公司)。测序由上海生工生物技术公司完成。
1.2 方法
1.2.1引物设计与合成 本研究所用引物名称、序列及用途见表1。引物合成由上海生工生物技术公司完成。

表1 引物序列表
1.2.2CFT073iucA基因缺失株的构建 外源性打靶片段的扩增与纯化:以pKD3质粒为模板,iucA-F和iucA-R作为引物,扩增含氯霉素的打靶基因。
Red重组系统的诱导表达和外源线性打靶片段的电转化:制备大肠埃希菌CFT073菌株的电转化感受态细胞。将pKD46质粒电转化入CFT073细胞,涂布于氨苄青霉素(100 μg/ml)平板,30℃培养过夜。挑选CFT073/pKD46单克隆,制备成电转化感受态细胞。iucA打靶片段转化进入CFT073/pKD46感受态细胞。转化细胞涂布于氯霉素(chloramphenicol,Cm)平板(含34 μg/ml Cm),37℃培养过夜。随机挑选22个克隆,接种LB液体培养基 (含34 μg/ml Cm),37℃培养过夜。用基因组同源手臂外侧的iucA-outF和iucA-outR引物进行PCR检测,当iucA基因被Cm抗性基因替换后,引物的扩增产物长度由1 921 bp缩短为1 292 bp。选择替换成功的阳性克隆,命名为CFT073/ΔiucA-Cm。
CFT073/ΔiucA-Cm克隆中Cm抗性基因的删除:制备CFT073/ΔiucA-Cm的电转化感受态细胞,将pCP20质粒电转化入上述细胞。涂布于LB平板(含100 μg/ml氨苄青霉素),30℃培养过夜。挑选单克隆划线接种LB平板(无抗性),42℃培养过夜。随机挑取7个克隆,分别接种LB培养基,37 ℃培养过夜。各取0.5 μl菌液用iucA-outF和iucA-outR引物再次进行PCR扩增鉴定,Cm抗性基因消除的克隆产物长度缩短为359 bp。选择阳性克隆的PCR产物进行测序验证,iucA被成功敲除,命名为ΔiucA。
1.2.3CFT073iucA基因回补株的构建 采用携带四环素抗性基因的pRK415质粒作为大肠埃希菌的回补质粒。利用iucA-comF和iucA-comR引物从大肠埃希菌CFT073原始菌株中扩增iucA基因,经HindIII和EcoRI酶切后克隆入pRK415相应位点,得到回补质粒pRK415-iucA。制备ΔiucA电转化感受态细胞。通过电转化将pRK415-iucA回补质粒转化入ΔiucA菌株,在四环素平板上筛选阳性克隆。经PCR和测序验证,确定回补质粒转入目标菌株,选定其中一个克隆为iucA基因回补菌株,命名为C-iucA。
1.2.4CFT073、ΔiucA、C-iucA增殖曲线的测定 CFT073、ΔiucA和C-iucA菌株在LB培养液和无菌尿液中增殖曲线的测定方法见本课题组既往发表的研究[9]。
1.2.5体外黏附、侵袭实验 冻存的细胞复苏后至少传代两次,将5637细胞接种至24孔板,待长满至细胞形成单层。按照细菌:细胞为100:1的比例将细菌接种到单层细胞。37 ℃,5% CO2孵育2 h。黏附实验中,孵育2 h后,用PBS清洗5遍,加入0.5%Triton X-100 裂解细胞,吸出每孔样品梯度稀释涂板计数,作为黏附到细胞的细菌数。侵袭实验中,每孔加入终浓度为200 mg/L的庆大霉素孵育杀死细胞外细菌,PBS清洗3遍,加入0.5%Triton X-100 裂解细胞,吸出每孔样品梯度稀释涂板计数,作为侵袭到细胞内的细菌数。黏附率和侵袭率计算公式如下:
黏附率=黏附到细胞的细菌数/接种总菌数×100%
侵袭率=侵袭到细胞内的细菌数/接种总菌数×100%
1.2.6体内定植能力检测 C57B/L6小鼠尿路感染模型的建立方法及CFT073、ΔiucA和C-iucA菌株在膀胱定植能力检测见本课题组既往发表的研究[10]。
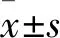
2 结果
2.1 CFT073、ΔiucA和C-iucA菌株增殖曲线测定将CFT073、ΔiucA和C-iucA菌株分别接种于LB液体培养基和无菌尿液后,每隔2 h取样测定A600,绘制增殖曲线显示,在LB液体培养基中CFT073、ΔiucA和C-iucA菌株具有相似的增殖速率(F=2.613,P=0.153)(图1)。而在无菌尿液中,三者增殖速率差异有统计学意义(F=18.927,P=0.003),其中ΔiucA的增殖速率低于CFT073菌株(P=0.001),C-iucA的增殖速率则高于ΔiucA(P=0.005),C-iucA与CFT073菌株增殖速率比较,差异无统计学意义。见图2。

图1 CFT073、ΔiucA和C-iucA菌株在LB液体培养基中的增殖曲线
2.2 体外黏附和侵袭能力比较比较CFT073、ΔiucA、C-iucA菌株对人膀胱癌上皮细胞株5637的黏附率和侵袭率显示,ΔiucA组黏附率和侵袭率均低于CFT073组(P=0.007、 0.002)(图3、4)。而C-iucA组黏附率和侵袭率与ΔiucA组相比均有所上升(P=0.046、 0.037)(图3、4)。C-iucA组黏附率接近CFT073组,两组之间差异无统计学意义(P=0.177)(图3),侵袭率则仍低于CFT073组,差异有统计学意义(P=0.040)(图4)。
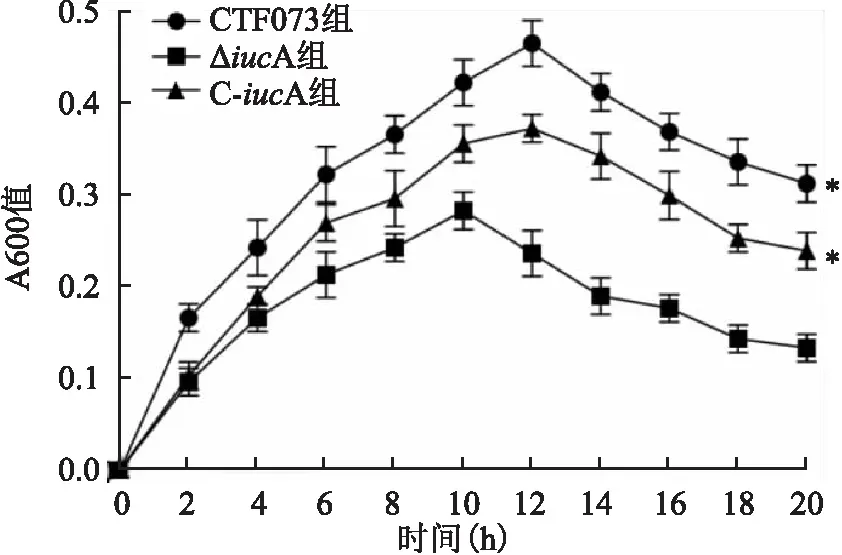
图2 CFT073、ΔiucA和C-iucA菌株在无菌尿液中的增殖曲线与ΔiucA组比较:*P<0.05
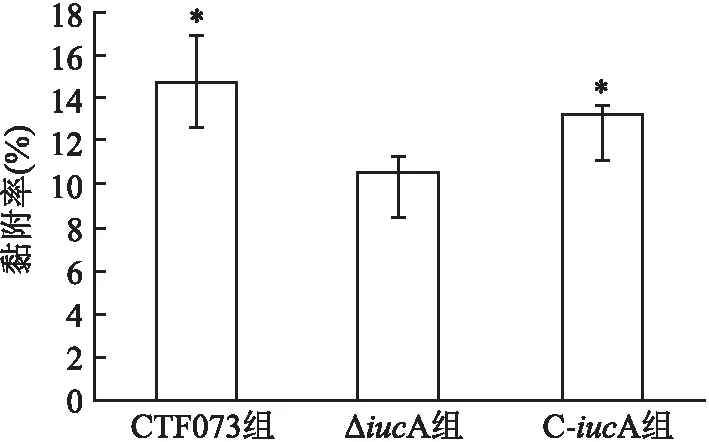
图3 CFT073、ΔiucA和C-iucA菌株对人膀胱癌上皮细胞5637的黏附率与ΔiucA组比较:*P<0.05
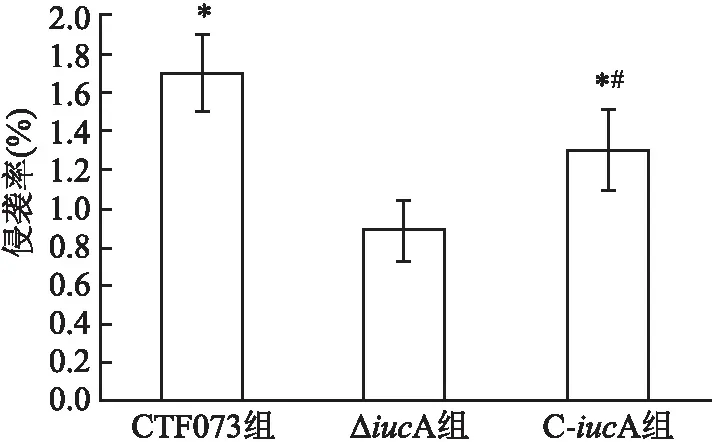
图4 CFT073、ΔiucA和C-iucA菌株对人膀胱癌上皮细胞5637的侵袭率与ΔiucA组比较:*P<0.05;与CFT073组比较:#P<0.05
2.3 膀胱定植能力检测成功构建C57B/L6小鼠UTI模型后,CFT073、ΔiucA和C-iucA菌株通过尿道灌注的方法感染小鼠,取膀胱组织匀浆涂平板培养计算定植细菌数。采用多个独立样本比较的秩和检验分析显示,CFT073组侵袭到膀胱定植的细菌数平均秩次为17.75,ΔiucA组为6.00,C-iucA组为13.75,各组之间定植细菌数差异有统计学意义(Kruskal-Wallis秩和检验卡方值为11.545,P=0.003)。各组之间两两比较显示,ΔiucA组与CFT073组,ΔiucA组与C-iucA组之间膀胱定植细菌数比较差异均有统计学意义(P=0.002、0.017), CFT073组与C-iucA组相比,定植细菌数差异无统计学意义。见图5。

图5 CFT073、ΔiucA和C-iucA菌株在小鼠膀胱的定植细菌数与ΔiucA组比较:*P<0.05
3 讨论
铁载体作为致病过程中重要的毒力因子和可能的治疗靶标已经在多种致病菌中被证实[11]。本课题组既往研究已经证实铁摄取相关基因fyuA在UPEC致病中的作用[9-10]。本研究继续探究铁载体气杆菌素编码基因iucA与UPEC致病之间的关系。
既往研究中,Rosso et al[12]通过分别构建高毒力肺炎克雷伯菌(hypervirulentKlebsiellapneumoniae, hvKP)的气杆菌素、沙门菌素、肠杆菌素和耶尔森杆菌素等铁载体的合成缺失株,发现与野生株相比,hvKP1ΔiucA菌株(气杆菌素合成缺失株)在腹水和血清中的存活受到影响,生长速率减低,并且在小鼠动物模型中hvKP1ΔiucA菌株表现为毒力减低。而沙门菌素、肠杆菌素和耶尔森杆菌素的单独或联合合成缺失株均不影响菌株在体外生长/存活以及在小鼠体内毒力。提示iucA影响菌株生长繁殖,是hvKP的关键毒力因子。Khasheii et al[13]的研究也表明,与合成其他铁载体的菌株相比,合成耶尔森杆菌素和气杆菌素的UPEC表现出最高的生长速率。提示气杆菌素可能有助于菌株的生长繁殖。本研究也显示,尽管在LB液体培养基中野生株、缺失株和回补株表现为相似的增殖曲线,但是在无菌尿液中,iucA缺失株的生长速率低于野生株,而iucA基因的回补显著回复菌株的生长繁殖能力,提示当UPEC处于铁含量较低的环境(无菌尿液)时,iucA基因对促进UPEC生长繁殖具有重要作用。推测iucA基因这种对菌株生长繁殖的促进作用与其促进铁离子的摄取作用有关,当处于营养丰富的环境(LB培养液)中时,尽管缺失了iucA基因,不能合成气杆菌素,UPEC仍可以通过其它铁载体介导铁离子的摄取满足菌株生长繁殖所需,而当UPEC处于铁含量较低的无菌尿液中时,iucA基因的缺失进一步削弱了菌株对铁离子的利用能力,导致生长繁殖受限。其具体机制尚需要进一步实验证实。
黏附和侵袭是病原菌致病的第一步,强大的黏附能力可以帮助尿路致病菌抵抗尿液的冲刷,黏附在上皮细胞后再侵袭进入胞内发挥致病作用。在本研究中,通过野生株、缺失株和回补株分别与人膀胱癌上皮细胞株5637相互作用显示,iucA缺失株的黏附率和侵袭率均低于野生株, 而在回补了iucA基因后,黏附率和侵袭率与缺失株相比均有所上升。提示iucA基因在促进菌株黏附和侵袭方面同样具有重要作用。为进一步明确iucA基因这种介导菌株黏附和侵袭的作用是否有助于菌株在靶器官的定植,课题组建立了小鼠尿路感染的动物模型。3种菌株分别感染C57B/L6小鼠后,缺失株在小鼠膀胱的定植数目低于野生株,而回补株可以回复其在膀胱组织的定植能力,提示iucA基因有助于增强UPEC在靶器官的定植能力。这与Ling et al[8]发现缺失iucA基因的禽致病性大肠埃希菌在肝脏、肾脏、脾脏、心脏和肺脏等的定植能力大幅减低的研究结论相似。iucA基因增强UPEC在靶器官的定植能力的机制,可能部分与其促进菌株的生长繁殖以及增加菌株的黏附和侵袭能力有关。
本研究通过iucA基因的缺失和回补,明确了铁载体气杆菌素在促进UPEC生长繁殖、体外黏附和侵袭以及体内靶器官定植中的作用。这有助于筛选与UPEC致病密切相关的铁载体,可以为UTI抗菌药物作用靶点、疫苗候选抗原选择奠定基础,并且对其他肠杆菌科细菌的致病机制研究具有借鉴意义。iucA基因介导UPEC生长、黏附和侵袭的具体作用机制未来还需要通过进一步的实验研究明确。
