基于宏基因组技术分析固态发酵枣酒酒醅的微生物多样性及关键风味基因
2022-02-16夏亚男
夏亚男,双 全*
(1.内蒙古农业大学 乳品生物技术与工程教育部重点实验室, 内蒙古 呼和浩特 010000;2.内蒙古农业大学食品科学与工程学院,内蒙古 呼和浩特 010000)
枣酒作为一种独具特色的水果酒,是以红枣为原料,经固态发酵酿造而成。早在西周时期,为了保存红枣,人们就想出了一种利用红枣发酵酿造枣酒的办法。枣酒不仅风味独特、枣香浓郁,而且酒性柔和、营养丰富。红枣中所含的芦丁、多糖和其他对人体健康有益的物质在发酵过程中被转化和水解,从而大大提高了其溶解度和吸收率,营养价值也相应提高[1-2]。但枣酒的生产主要是基于小作坊模式,未形成标准化的生产工艺及商业化发酵剂,加之产品质量的不稳定性,限制了枣酒产业的发展,也影响了红枣资源的加工利用和资源优势的转化[3-4]。因此,加深对枣酒发酵的科学研究,能够为枣酒产区充分发挥资源优势,逐渐形成红枣区域性特色支柱产业具有重要的现实意义。
固态发酵方式是我国白酒的一种独特发酵模式,它比液态发酵会形成更加丰富的微生物菌系,从而引起更加复杂的微生物代谢活动,所形成的代谢产物,如风味也会比液态发酵更加浓郁、醇厚[5-6]。而相关研究也主要集中在白酒酒曲、酒醅及窖泥的成分及微生物菌群结构研究[7-9],如胡晓龙等[10]采用高通量测序技术在中温大曲中检测到了28个属,其中肠杆菌属、乳杆菌属为大曲样品中的优势细菌属。截至目前,缺少对水果类原料固态发酵酒酒醅的相关研究,无法为固态发酵红枣蒸馏酒的生产及工艺优化提供指导,而且对白酒酒曲、窖泥等的微生物菌群结构研究大多采用的是16S rRNA高通量测序技术,只能确定到微生物的属分类水平,不能确定具体的菌种种类,从而无法明确地为特定菌株筛选、风味形成及功能特性研究提供指导。
随着现代分子生物学技术的发展,宏基因组技术开始逐渐应用于食品微生物的研究中。宏基因组学是一种基于基因序列分析环境样品中微生物信息的研究方法[11-12]。它可以快速、准确地获取大量的生物学数据和丰富的微生物信息,已成为研究微生物多样性和群落特征的重要手段。相比于16S rRNA高通量测序技术,宏基因组技术不仅可以全面地挖掘出微生物基因组信息,而且能够进行功能及代谢通路的分析,基于这些信息,更便于挖掘菌群的生物多样性、群落结构、功能特性、相互关系[13-15]。因此,本实验基于宏基因组技术考察固态发酵枣酒酒醅的微生物菌群多样性,分析其主要代谢通路,并挖掘与风味形成相关的关键功能基因,以期为我国枣酒的品质提升及风味功能基因的利用提供理论依据。
1 材料与方法
1.1 材料、试剂与仪器
酒醅取自河北省阜平县董家枣酒厂。原料枣为河北沧州小枣。酒醅制取:取去核红枣,按枣∶水=1∶1比例浸泡、粉碎后,按1%接种酒曲,在自然窖池中26~28 ℃发酵14 d,获得固态发酵枣酒酒醅,-80 ℃冷冻,用于后续实验。
E.Z.N.A.®Soil DNA Kit 美国Omega Biotek公司;琼脂糖 西班牙Biowest公司;FastPfuPolymerase 北京全式金生物技术有限公司;AxyPrep DNA Gel Extraction Kit美国Axygen公司。
NanoDrop2000超微量分光光度计 美国Thermo Fisher Scientific公司;DYY-6C电泳仪 北京市六一仪器厂;GeneAmp®9700型聚合酶链式反应(polymerase chain reaction,PCR)仪 美国ABI公司;5424R高速台式冷冻离心机 美国Eppendorf公司;MiSeq PE300测序仪 美国Illumina公司。
1.2 方法
1.2.1 酒醅样品DNA抽提
E.Z.N.A.®Soil DNA Kit试剂盒用于提取样品微生物的宏基因组DNA。基因组DNA的提取完成后,通过1%琼脂糖凝胶电泳检测提取的基因组DNA。
1.2.2 宏基因组检测
构造PE库:连接“Y”形接头;使用磁珠去除接头自身连接的片段;使用PCR扩增以丰富文库模板;使氢氧化钠变性以生成单链DNA片段。
桥式PCR:DNA片段的一端与引物碱基互补并固定在芯片上;另一端与附近的另一个引物随机互补,并且也固定并形成“桥”。PCR扩增产生DNA簇;DNA扩增子线性化为单链。
Illumina HiSeq测序:添加带有4 种荧光标记的修饰的dNTP和DNA聚合酶,每个循环仅合成一个碱基;用激光扫描反应板的表面,读取每个模板序列第一轮反应聚合的核苷酸种类;化学切割“荧光基团”和“终止基团”,恢复3’末端黏性,并继续聚合第2个核苷酸;计算每一轮收集的荧光信号的结果,并获得模板DNA的序列。
1.2.3 生物信息分析
序列拼接组装:使用Megahit拼接软件对clean序列进行拼接组装,根据不同的kmer大小进行组装,从中选择最优组装结果。
基因预测:使用MetaGeneMark(http://topaz.gatech.edu/GeneMark/meta_gmhmmp.cgi)对拼接结果中的contig进行基因预测,并将其翻译为氨基酸序列。
非冗余基因集的构建:将所有样品预测出的基因序列,用CD-HIT软件进行聚类,构建非冗余基因集。将样品所有的高质量reads与非冗余基因集进行比对,统计基因在对应样品中的丰度信息,从而构建geneprofile。比对软件:BWA(http://bio-bwa.sourceforge.net/)。
宏基因组物种注释及丰度分析:通过Kraken软件对测序样品数据进行物种注释,获得物种分类及丰度信息。
直系同源群集(Cluster of Orthologous Group,COG)功能注释:通过与COG数据库进行BLASTp比对,可以获得基因所对应的COG注释,即得出基因对应的COG数,并根据COG数进行功能归类。
京都基因与基因组百科全书(Kyoto Encyclopedia of Genes and Genomes,KEGG)注释:运用FMAP软件对获得基因进行KEGG注释和差异Module分析。
碳水化合物活性酶注释:通过将基因序列与CAZy(Carbohydrate-Active Enzymes)数据库进行比对,可以获得碳水化合物酶类的物种来源、酶功能EC分类、基因序列、蛋白质序列及其结构等信息。
1.3 数据处理
采用R软件进行统计学分析及作图。
2 结果与分析
2.1 酒醅微生物多样性分析
枣酒酒醅样品中基因组全长128 785 299 bp,包含356 189 条序列,G+C含量为43.46%。对枣酒酒醅基因进行物种丰度注释,共鉴定出微生物37个门、1 247个属、3 937个种,其中相对含量大于1%的优势菌群在门、属、种水平的数量分别为4、5、10 种(图1)。
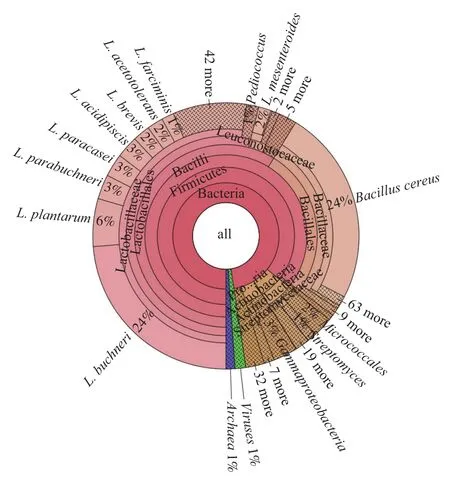
图1 枣酒酒醅样品微生物群落Circos图Fig. 1 Circos diagram of microbial community in fermented jujube mash
在微生物门水平上,优势菌门有厚壁菌门(Firmicutes)、放线菌门(Actinobacteria)、变形菌门(Proteobacteria)、广古菌门(Euryararchaeota)。其中厚壁菌门(84.60%)为绝对优势菌门,存在较高的相对丰度。这与严超[16]研究的红枣白兰地酒醅中优势细菌门类似。在属水平上,优势菌属包括乳杆菌属(Lactobacillus)、芽孢杆菌属(Bacillus)、明串珠菌属(Leuconos)、片球菌属(Pediococcus)和链霉菌属(Streptomyces)。其中乳杆菌属(53.24%)和芽孢杆菌属(20.51%)含量最高。此结果与袁帅[17]研究的茅台酒醅和国台酒醅中优势菌群相似,其中乳酸菌属都是优势菌群。
在种分类水平上,相对含量大于1%的优势菌种有10 个,包括布氏乳杆菌(L. buchneri)、蜡样芽孢杆菌(Bacillus cereus)、植物乳杆菌(L. plantarum)、类布氏乳杆菌(L. parabuchneri)、副干酪乳杆(L. paracasei)、嗜酸乳杆菌(L. acidipiscis)、短乳杆菌(L. brevis)、耐酸乳杆菌(L. acetotolerans)、肠膜明串珠菌(Leuconostoc mesenteroides)、香肠乳杆菌(L. farciminis),相对含量分别为24%、24%、6%、3%、3%、3%、2%、2%、2%、1%。排名前10的菌种中9种属于乳酸菌,其中8 种是乳杆菌,可见乳杆菌是固态发酵枣酒酒醅的绝对优势菌种。
L. buchneri是一种异型发酵的乳酸菌,在发酵过程中能将乳酸分解成乙酸和丙二醇,而乙酸等挥发性脂肪酸是比较有效的抗真菌及霉菌的酸类物质,能够适当抑制杂菌的生长,维持发酵菌的正常生长、代谢等活动[18]。蜡样芽孢杆菌主要存在于土壤、水、空气以及动物肠道等处,可产生抗菌物质,抑制有害微生物的繁殖,降解土壤中的营养成分,改善生态环境,也会因食品贮藏不当而导致轻微食物中毒状况。酒醅中蜡样芽孢杆菌的存在与窖池泥土的自然发酵环境有关。而固态发酵环境普遍比液态发酵模式具备更丰富的微生物菌系,能酿造出更加浓郁的风味。如蜡样芽孢杆菌产中性蛋白酶、磷脂酶等的能力较强,有助于风味物质等代谢产物的生成[19-20]。酒醅中尽管存在少量有害菌,但后期采用蒸醅取酒,获得酒醅的蒸馏液,并不直接食用酒醅,所以并未引起相应安全问题。母应春等[21]发现酒曲中的细菌优势菌门为变形菌门,优势细菌属为红球菌属;左乾程等[22]发现清酱香型白酒酒醅发酵过程中核心细菌属为Lactobacillus、Bacillus、Weissella、Acetobacter等,随发酵进行,Lactobacillus逐渐成为主导。目前鲜见采用宏基因组技术检测枣酒酒醅微生物多样性的报道,以往研究只确定到酒醅的属水平,该结果对微生物种水平的确定丰富了枣酒微生物多样性的具体类别信息。
枣酒发酵过程中产生的风味物质与酒醅中微生物的生长和代谢密切相关。这些微生物催化原料不断进行一系列复杂的反应,可以分解原料中的淀粉、蛋白质,形成糖和氨基酸,从而引发美拉德反应,生成枣酒独特的风味前体物质;能够将蛋白质分解为小分子肽,并形成重要的香气成分,例如醛、醇、酯、酮等,进而形成枣酒清爽醇厚的风格特征[23-25]。酒醅在酒类生产中扮演了极其重要的角色,其中所含微生物的数量和种类决定了成品酒的优劣。
2.2 常用数据库注释分析
2.2.1 COG数据库注释分析
将基因翻译获得的蛋白质序列与COG数据库进行比较,可获得相应的蛋白质功能注释信息(图2),酒醅样品中共鉴定到了6 269 个COG功能单元,这些功能单元分属于23个主要的功能大类,其中相对含量在前8 位的功能大类是氨基酸运输代谢,翻译、核糖体结构及生物合成,碳水化合物转运与代谢,转录,复制、重组与修饰,无机离子运输和代谢,能量产生与转换,细胞壁/膜/被膜生物起源,基因数分别为746、702、660、520、449、395、388、384个。除微生物基本生命活动外,代谢活动主要以氨基酸代谢和碳水化合物代谢为主。本研究与Chen Chen等[26]采用宏基因组分析酒曲中微生物多样性与风味形成相关性的结果一致,在酒曲所有活性代谢途径中,膜转运、碳水化合物代谢和氨基酸代谢最为活跃,而碳水化合物代谢和氨基酸代谢这两类代谢通路与风味物质形成密切相关。
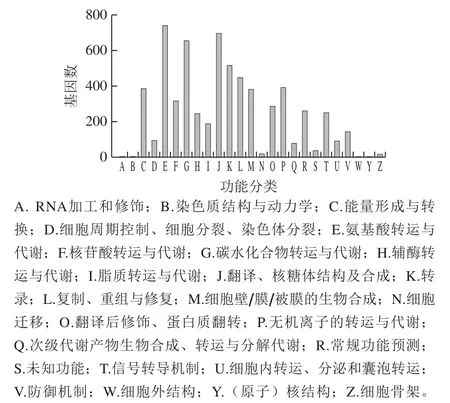
图2 枣酒酒醅基因COG功能分类Fig. 2 COG Functional classification of genes in fermented jujube mash
2.2.2 KEGG数据库注释分析
枣酒酒醅中238 860 个不同的基因被注释,从属于KEGG数据库6大类 43个KEGG 通路中(图3)。6大类分别是代谢通路(133 173,55.75%)、环境信息处理(29 448,12.33%)、遗传信息处理(27 570,11.54%)、细胞进程(18 558,7.77%)、人类疾病(18 291,7.66%)和生命系统(11 820,4.95%)。富集差异表达基因最多的8个KEGG通路包括碳水化合物代谢(38 526)、氨基酸代谢(22 671)、膜运输(17 493)、辅酶和维生素代谢(14 232)、能量代谢(14 022)、核苷酸代谢(11 979)、信号传导(11 910)、翻译(11 457)。其中,碳水化合物代谢及氨基酸代谢是最主要的两类代谢活动,与COG功能注释结果一致。Chen Chen等[26]利用Spearman相关性分析了特定代谢通路与风味形成的关系,发现根霉(Rhizopus)、酵母菌(Saccharomycopsis)、Wickerhamomyces和Weissella菌种与氨基酸代谢呈正相关,而碳水化合物代谢与Rhizopus、Saccharomycopsis、Pediococcus和Weissella有关。
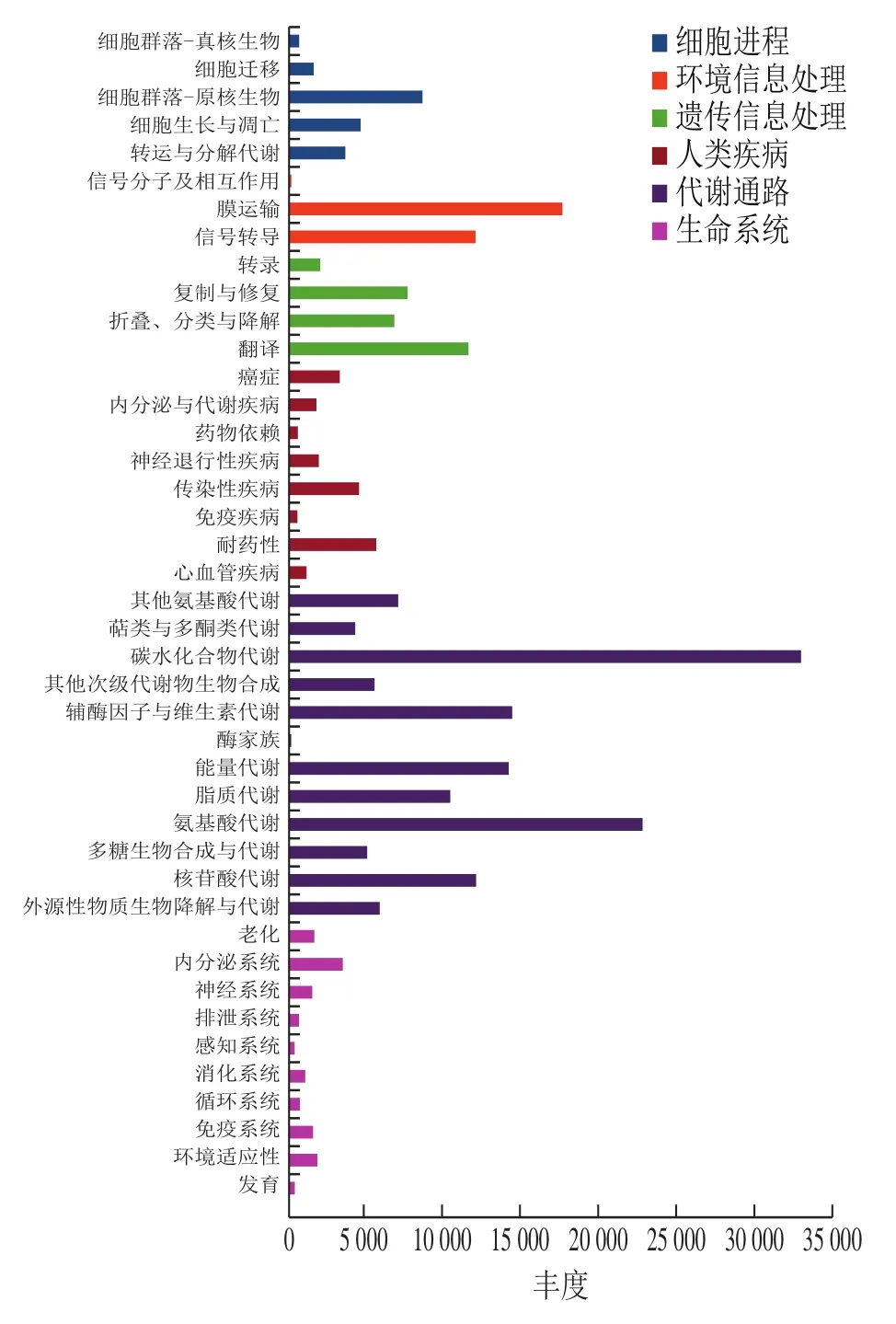
图3 枣酒酒醅基因KEGG代谢通路统计Fig. 3 Statistics of KEGG metabolic pathways in fermented jujube mash
2.2.3 基于CAZy数据库的碳水化合物活性酶分析
CAZy数据库可将不同种类的碳水化合物活性酶分为糖苷水解酶、糖基转移酶、多糖裂合酶、碳水化合物酯酶、碳水化合物结合模块、辅助模块酶类6大类蛋白质家族。与CAZy数据库比对后,枣酒酒醅样品中共鉴定出3 824个碳水化合物活性酶(图4),其中,糖苷水解酶(1 232个)和糖基转移酶(1 238个)类数量最多,占据样品检测数据总数的70%。其次是碳水化合物酯酶(533个)、碳水化合物结合模块(416个)和辅助模块酶类(157个)。多糖裂合酶(48个)类最少,仅占样品检测到酶总数的0.9%。糖苷水解酶以内切或外切方式水解寡糖、多糖等各种含糖化合物的糖苷键,生成单糖、寡糖或糖复合物,因而在寡糖、芳香基糖苷的合成、氨基酸和多肽的糖基化方面发挥了重要作用。而糖基转移酶在生物体内催化活化的糖连接到不同的受体分子,如蛋白、寡糖、脂和小分子上[27-29]。酒醅中丰富的糖苷水解酶及糖基转移酶为单糖、寡糖的形成、转移及进一步代谢提供了基础。
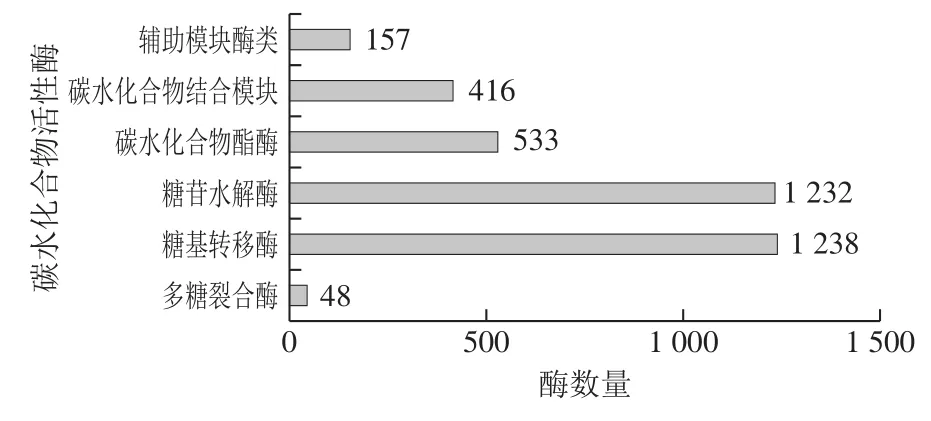
图4 枣酒酒醅CAZy统计Fig. 4 Statistics of CAZy in fermented jujube mash
2.3 糖代谢基因分析
2.3.1 糖转运系统
糖转运系统中,甘露醇、果糖、甘露糖和蔗糖可同时由磷酸葡萄糖转移酶系统(glucose phosphotransferase system,PTS)转运,由表1可知,枣酒酒醅中分别发现控制基因20、17、200、28个,表明发酵过程中酒醅微生物对不同糖的转运能力不同,发酵期间菌种对甘露糖的转运能力较强。乳糖和麦芽糖可以通过PTS、ABC(ATP binding cassette)转运蛋白以及透性酶3种方式进行转运。但是酒醅中未发现关于麦芽糖的PTS及透性酶基因,ABC转运蛋白中有多糖转运和单糖转运蛋白两类,未发现麦芽糖的特异性转运系统基因,表明酒醅中微生物对麦芽糖的转运能力较弱,较难实现胞外分解。半乳糖醇只能通过PTS进行转运,酒醅中发现半乳糖醇转运蛋白相关基因79 个。葡萄糖酸盐、果糖和低聚糖也可利用相关透性蛋白进入菌株细胞内,分别检索到基因2、8、35个。

表1 酒醅糖转运系统基因分析Table 1 Gene analysis of carbohydrate transport systems in fermented jujube mash
2.3.2D-1-磷酸果糖的转化
果糖经PTS转运最终进入细胞后形成D-1-磷酸果糖,而后转化为糖酵解的中间产物D-3-磷酸甘油醛。此过程可通过3 条途径实现,关键酶分别是磷酸果糖激酶和2-磷酸果糖醛缩酶,二磷酸果糖醛缩酶和磷酸丙糖异构酶,以及2-磷酸果糖醛缩酶和丙糖激酶3种组合。结合表1、2,酒醅中含有果糖的PTS,且含有11个二磷酸果糖醛缩酶和32个磷酸丙糖异构酶基因,可通过第2种途径将胞内的D-1-磷酸果糖转化为D-3-磷酸甘油醛。
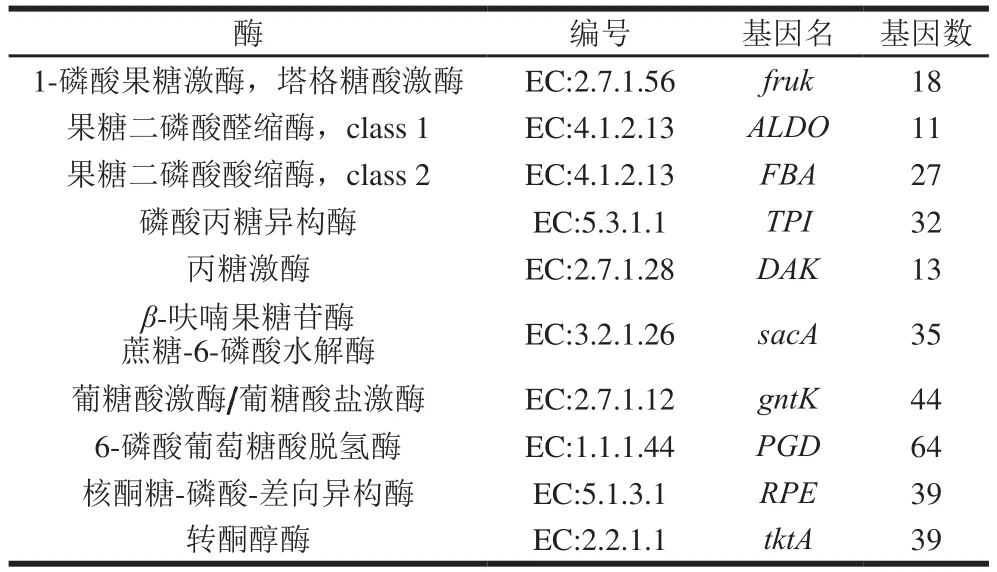
表2 酒醅中参与糖类转化为糖酵解中间产物的关键酶基因分析Table 2 Genes encoding enzymes involved in the conversion of carbohydrates into intermediates in glycolysis pathway
2.3.3 6-磷酸蔗糖的转化
蔗糖经PTS转运至细胞后形成6-磷酸蔗糖,之后可在β-呋喃果糖苷酶/6-磷酸蔗糖水解酶的催化作用下转化为D-6-磷酸葡萄糖和β-D-果糖。结合表1、2,酒醅中含有果糖的PTS,且编码了35个6-磷酸蔗糖水解酶的基因,因此酒醅可以利用胞外蔗糖,转运至胞内后,进一步生成D-6-磷酸葡萄糖和β-D-果糖。
2.3.4 葡萄糖酸盐的转化
转运至细胞内的葡萄糖酸盐可进一步形成糖酵解途径的中间物质β-D-6-磷酸果糖,但此过程需要在葡糖酸激酶、6-磷酸葡萄糖酸盐脱氢酶、磷酸核酮糖-3-差向异构酶和转酮醇酶的催化作用下才能实现。酒醅中发现4 种酶的控制基因,基因数分别为44、64、39、39 个,因此酒醅在发酵过程中可利用胞外的葡萄糖酸盐,进一步生成β-D-6-磷酸果糖,参与糖酵解途径。
2.4 氨基酸风味形成途径的基因分析
氨基酸风味形成途径起始于转氨作用,氨基酸经转氨酶转化为相应的酮酸,酮酸较不稳定,转化成相应的醛类,进一步脱氢生成醇类,再经酯化转化为相应的酯类,这些代谢物都是食品中重要的风味成分。
2.4.1 转氨酶
支链氨基酸、芳香族氨基酸和蛋氨酸的转氨作用可以通过不同的转氨酶催化。支链氨基转移酶(branchedchain aminoacid transferase,BcAT)对支链氨基酸和蛋氨酸具有活性。如表3所示,在枣酒酒醅基因中,发现BcAT基因(ilvE)171个,其可以催化支链氨基酸。例如,支链氨基酸中的亮氨酸、异亮氨酸和缬氨酸可通过支链氨基转氨酶分别转化为一系列α-酮异己酸、α-酮基-β-甲基戊酸和α-酮基-β-甲基丁酸。其中,据报道α-酮异己酸是发酵香肠中的主要风味物质。在枣酒中,诸如2-甲基丁酸乙酯(汗味)和异丁酸(酸味和甜味)之类的风味化合物很可能是通过异亮氨酸或缬氨酸在转氨酶作用下经氨基酸转氨作用生成。
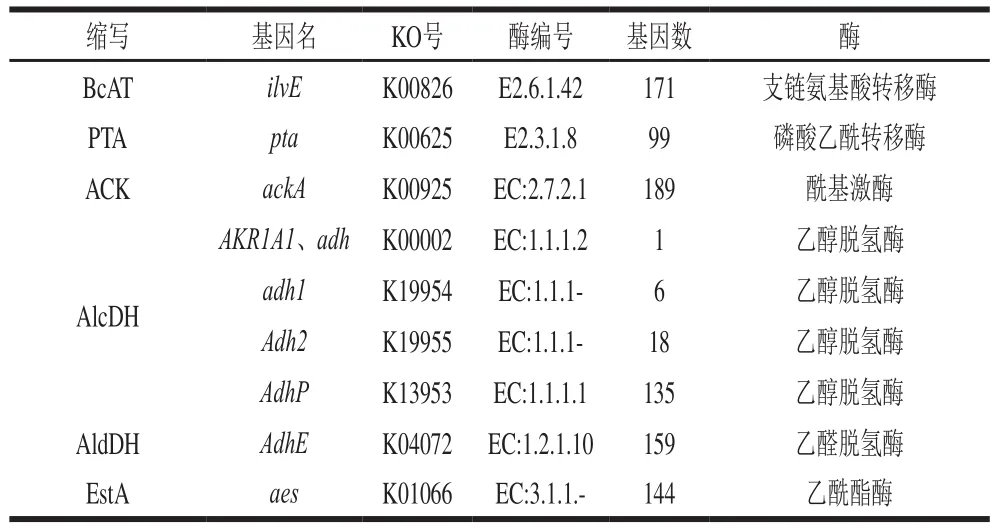
表3 枣酒酒醅基因组中参与转氨作用途径的酶编码基因Table 3 Genes encoding enzymes involved in transamination pathway in fermented jujube mash
2.4.2 酮酸转化酶
酮酸可以3种方式进行转化。第1种方式:酮酸通过氧化脱羧直接转化为羧酸,主要依靠酮酸脱氢酶(ketoacid dehydrogenase,KaDH)、磷酸转乙酰酶(phosphotransacetylase,PTA)和酰基激酶(acyl kinase,ACK)。如表3所示,在枣酒酒醅中存在99 个PTA(pta)和189 个编码ACK(ackA)基因,表明酒醅发酵过程中可以通过该途径将支链氨基酸(亮氨酸、异亮氨酸和缬氨酸等)的酮酸转化为甜味的甲基丁酸和异丁酸。第2种方式是通过羧酸脱氢酶将酮酸分解为羧酸。第3种方式是α-酮酸通过α-酮酸脱羧酶转化为相应的醛。在枣酒酒醅中未发现编码羧酸脱氢酶和酮酸脱羧酶的基因,这表明酒醅中的微生物无法进行后两种转化形式,发酵过程中主要采用第1种酮酸转化方式。
2.4.3 醇和醛脱氢酶
醛脱氢酶和醇脱氢酶可以分别催化醛(醇)类物质转化成相应的醇类和羧酸类物质,控制基因分别为醛脱氢酶(aldehyde dehydrogenase,AldDH)和醇脱氢酶(alcohol dehydrogenase,AlcDH)。枣酒酒醅中发现159个编码AldDH的基因(AdhE),因此酒醅中的微生物具有催化醛转化为相应醇的潜力。例如,它可以催化异戊醛、活性戊醛和异丁醛以生成相应的异戊醇、活性戊醇和异丁醇。适量的高级醇给枣酒带来丰厚的醇和感,使其绵柔利口,此结果与枣酒中丰富的高级醇含量相一致[30-31]。同时,酒醅中发现160 个编码AlcDH的基因,分为adh、adh1、Adh2、AdhP4 种基因类型,其中AdhP(135)基因数最丰富,表明酒醅中微生物可将醇类转化为相应的羧酸类物质。
3 结 论
传统枣酒香气浓郁、风味独特,与其酒醅中复杂的微生物代谢活动密不可分。本实验基于宏基因组技术分析枣酒酒醅的微生物多样性,挖掘糖代谢及氨基酸风味形成系统的功能基因。主要结论如下:
1)酒醅样品共检测到356 189 条序列,G+C含量为43.46%。对枣酒酒醅基因进行物种丰度注释,共鉴定出微生物37个门、1 247个属、3 937个种,其中相对含量大于1%的优势菌群在门、属、种水平的数量分别为4、5、10 个;核心菌种是L. buchneri、L. plantarum、L. parabuchneri等。
2)经COG、KEGG数据库比对,分别注释到6 269、238 860 个基因,主要代谢活动为氨基酸代谢和碳水化合物代谢。碳水化合物活性酶中,糖苷水解酶(1 232)和糖基转移酶(1 238)数量最多,占据枣酒酒醅碳水化合物活性酶的70%。
3)在糖代谢途径中,酒醅编码了甘露醇、果糖、甘露糖、蔗糖和半乳糖醇的糖转运系统基因,检索到葡萄糖酸盐、果糖和低聚糖相关透性蛋白基因,且具备催化胞内D-1-磷酸果糖、6-磷酸蔗糖及葡萄糖酸盐的关键酶基因,具备将其转化为糖酵解中间产物的基因基础。
4)氨基酸风味形成中,酒醅编码了171个BcAT、288个酮酸转化酶、319 个醇/醛脱氢酶和144个乙酰酯酶的控制基因,氨基酸可经转氨酶转化为相应酮酸,进而转化成相应的醛类,进一步脱氢生成醇类,再经酯化转化为相应酯类,具备形成浓郁酯香风味的基础。
本实验在分析枣酒酒醅菌群结构组成的基础上,进一步明确了酒醅主要代谢活动及糖代谢、氨基酸代谢的关键酶基因,后续需进一步对优势菌株进行筛选应用,分析优势菌株在主要代谢通路中的作用,并进一步验证关键酶基因的表达情况,以期为枣酒的品质提升及微生物功能基因库的挖掘提供理论依据。
