小麦WRKY转录因子的鉴定及其在胚性愈伤组织形成中的表达分析
2022-01-09楚宗丽张睿男李亮杰孙君艳王付娟仝胜利
楚宗丽,张睿男,2,李亮杰,孙君艳,王付娟,周 强,仝胜利
(1. 信阳农林学院,河南信阳 464006; 2.南京农业大学农学院,江苏南京 210095)
小麦(TriticumaestivumL.)是我国重要的粮食作物之一,高产优质小麦品种的选育对保证国家粮食安全和社会稳定具有重要意义。生物技术为作物新品种培育提供了有效途径,但小麦组培效率较低,制约着生物技术在小麦育种中的应用。胚性愈伤组织容易再生获得植株,是提高小麦组培效率的关键。胚性愈伤组织的形成是一个复杂的调控过程。植物胚性愈伤组织的形成受AGAMOUS-LIKE15(AGL15)[1-2]、 BABY BOOM[3-4]、LEC[5-6]等转录因子的影响,同时,逆境胁迫因子对植物胚性愈伤组织的形成也有影响[7-8]。
WRKY转录因子参与植物的多种生理代谢过程,是一类重要的转录因子家族,其N端含有高度保守的WRKY结构域,该结构域约由60个氨基酸残基组成,包含高度保守的WRKYGQK序列,同时C末端含有一个锌指结构,锌指结构有C-X4-5-C-X22-23-H-X1-H(C2H2)和C-X7-C-X23-H-X1-C(C2HC)两种类型[9]。根据所含WRKY结构域的数目和锌指结构类型,WRKY转录因子一般分为Ⅰ、Ⅱ和Ⅲ3组,其中,第Ⅰ组含有2个WRKY结构域和C2H2锌指结构;第Ⅱ组含有1个WRKY结构域和C2H2锌指结构;第Ⅲ组含有1个WRKY结构域和C2HC锌指结构[10]。WRKY转录因子通过与(T)(T)TGAC(C/T)序列(W-box)特异性结合,调节启动子中含有该元件相关基因的表达。目前,在拟南芥[11]、水稻[12]、棉花[13]、豆类[14-15]、葡萄[16]、菠萝[17]、月季[18]、茶树[19]等多种植物中已经鉴定出WRKY转录因子。在小麦中,Niu等[20]利用小麦EST数据库鉴定到43个TaWRKY基因,Okay等[21]和Gupta等[22]用逆境胁迫下的转录组数据也分别鉴定到160和107个TaWRKY基因。
WRKY转录因子参与植物的防御调控过程,包括响应多种生物和非生物胁迫[9,12,20-24],如月季WRKY转录因子基因RcWRKY41在月季抗灰霉病中发挥作用[18],棉花WRKY转录因子基因GhWRKY39-1在转基因烟草中可提高对病原体的防御能力,并且可调节盐胁迫耐受性[25]。小麦WRKY转录因子基因TaWRKY01和TaWRKY33可增强拟南芥抗旱和耐热的能力[21]。同时,WRKY转录因子参与植物的衰老、萌发和休眠以及胚的形成等过程[26-28]。
然而,目前在全基因组范围内进行小麦WRKY转录因子的鉴定及其在小麦胚性愈伤组织形成中的功能还未见报道。本课题组前期通过转录组测序分析了小麦胚性愈伤组织形成的分子机制,发现部分WRKY转录因子家族基因在胚性愈伤组织形成中上调表达[29],但仅在所测序转录组数据范围内进行了分析。小麦基因组序列的公布,为小麦WRKY转录因子家族成员的分析提供了基础。本研究以小麦基因组数据为基础,在全基因组范围内进行了小麦WRKY转录因子的鉴定、分类、系统进化树绘制、motif保守元件分析和基因结构分析,并对WRKY转录因子相关基因在小麦胚不同发育时期的胚性和非胚性愈伤组织中的表达模式进行分析,研究WRKY转录因子在小麦胚性愈伤组织形成中的作用,以期为研究小麦胚性愈伤组织形成的分子机理奠定基础。
1 材料与方法
1.1 注释基因组序列的获取
从Ensembl Plants数据库(http://plants.ensembl.org/)中获取小麦基因组数据。同时,从PlantTFDB数据库(http://planttfdb.cbi.pku.edu.cn/)中下载水稻和拟南芥WRKY蛋白序列,用于多序列比对和系统进化树的绘制。
1.2 小麦WRKY转录因子的鉴定
从Pfam网站((http://pfam.xfam.org/family/PF03106)下载WRKY结构域的隐马尔科夫模型文件,Pfam编号为PF03106。使用Hummer Search 3.0软件,在小麦基因组序列中通过隐马尔科夫模型比对,搜索小麦WRKY转录因子,使用Perl程序,筛选Hummer搜索结果,筛选条件为E-value<1e-3[18]。为了确保已知基因组信息中所有小麦WRKY转录因子不被遗漏,从NCBI网站(https://www.ncbi.nlm.nih.gov/)获取拟南芥WRKY蛋白序列,通过Blast比对,搜索小麦注释基因组序列中小麦WRKY转录因子。去除上述两种操作结果中的重复序列,对余下的序列用SMART数据库(http:// smart.embl.de/)和Pfam-CDD工具(https://www.ncbi.nlm.nih.gov/cdd/)进行蛋白结构预测,去除不含WRKY结构域的序列,保留确定的小麦WRKY转录因子序列。利用Perl程序对TaWRKY蛋白的分子量、等电点进行分析。使用MEGA 7.0软件,参照拟南芥WRKY蛋白分类方法[24]对鉴定出的TaWRKY蛋白进行分类。
1.3 小麦WRKY家族系统进化树的绘制
从PlantTFDB数据库中分别挑选21个具有代表性的拟南芥和粳稻WRKY蛋白序列,利用MEGA 7.0软件,与TaWRKY蛋白进行多序列比对,绘制系统进化树。算法为邻接法(neighbor-joining,NJ),模型为泊松模型,校验参数bootstrap为1 000[24]。
1.4 小麦WRKY蛋白保守基序与基因结构分析
在Bio-Linux环境下,使用MEME 4.12.0软件预测分析TaWRKY蛋白保守基序。具体参数为:搜索motif的总个数为10,motif长度范围为6~50。使用Perl程序获取TaWRKY基因在染色体上的外显子、CDS、UTR位置,使用在线工具GSDS 2.0(http://gsds.cbi.pku.edu.cn/)对TaWRKY基因结构进行分析,并绘制基因结构图。
1.5 小麦胚性愈伤组织中TaWRKY家族基因的差异表达分析
以小麦品种周麦18为材料,在其幼胚和成熟胚培养中,对来源于幼胚的胚性愈伤组织(培养3、6和15 d,分别用IME3、IME6和IME15表示)和成熟胚的非胚性愈伤组织(培养3、6和15d,分别用ME3、ME6和ME15表示)进行转录组测序,对转录组数据(ID:PRJNA353135,登录号:SRP093588)[29]进行分析,获得TaWRKY基因,并对不同转录组文库中的TaWRKY基因进行差异表达分析。
1.6 差异表达基因实时定量PCR验证

实时定量PCR反应在Bio-Rad CFX96TM实时定量系统上进行,PCR反应体系为25 μL,包括逆转录反应产物(cDNA溶液)2.0 μL、正、反向引物各1.0 μL(引物序列详见表1)、2× SYBR Premix Ex TaqTMII 12.5 μL、RNase游离水8.5 μL。PCR反应程序:95 ℃预变性30 s;95 ℃变性5 s、60 ℃退火30 s,72 ℃延伸30 s,40个循环。以Actin基因(GenBank: AB181991)为内参,所有反应设置3个生物学重复,通过琼脂糖凝胶电泳和溶解曲线分析验证每对引物的特异性。以ME3文库中的基因CT值作为参照,其表达水平设为1.0,使用2-△△Ct方法计算基因的相对表达量[30],结果以平均值±标准差表示。

表1 qRT-PCR所用引物
2 结果与分析
2.1 小麦WRKY转录因子的鉴定及分类
用Hmmer Search 3.0和NCBI Blast鉴定,分别获得312和69个候选小麦WRKY转录因子,删除两者的重复部分,余下序列通过SMART数据库和Pfam数据库进行验证,去除保守结构域或锌指结构域不完整的序列,最后确定270个小麦WRKY转录因子(TaWRKY01~TaWRKY270)。根据WRKY保守结构域数目和锌指结构域类型,将这些WRKY转录因子分为Ⅰ、Ⅱ和Ⅲ 3组,其中,第Ⅰ组WRKY转录因子(TaWRKY01~TaWRKY41)均含有2个WRKY结构域和1个C2H2锌指结构域,第Ⅱ组WRKY转录因子(TaWRKY42~TaWRKY186)均含有1个WRKY结构域和1个C2H2锌指结构域,第Ⅲ组WRKY转录因子(TaWRKY187~TaWRKY270)均含有1个WRKY结构域和1个C2HC锌指结构域,数量分别是41、145和84个。蛋白理化特性分析结果表明,TaWRKY蛋白氨基酸序列长度为135~1 482 aa,等电点为4.59~10.86,分子量为14 796.1~165 344.2。
2.2 WRKY转录因子家族的系统发育分析
从PlantTFDB数据库中分别选择21个具有代表性的完整的拟南芥和粳稻WRKY蛋白序列,与鉴定出的TaWRKY蛋白序列一起,利用MEGA 7.0进行多序列比对分析,绘制系统进化树,结果如图1所示。在I、II、III 3个分组中,TaWRKY蛋白主要分布在第II组中,根据基因结构和系统进化树关系,可进一步分为3个亚组:II-a、II-b和 II-c(图1),在系统进化树中,每个分组聚类为一个分支,组II中II-a、II-b和II-c分别聚类为一个分支,总计5个分支。
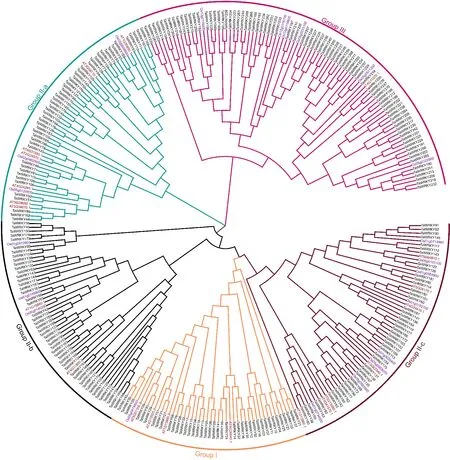
黑色、紫色和红色分别表示小麦、水稻和拟南芥中的WRKY成员。
2.3 TaWRKY蛋白保守基序分析
在Bio-Linux环境下,用MEME 4.12.0软件对TaWRKY蛋白序列进行保守基序扫描,共搜索到10个保守基序,命名为motif 1~motif 10(图2A),其中motif 1~ motif 5和motif 8均含有WRKY转录因子的WRKYGQK序列(如图2B),TaWRKY蛋白一般含有2~4个保守基序(除TaWRKY194、TaWRKY241、TaWRKY203和TaWRKY216仅含1个基序)(图2A),基序分布特征相似的TaWRKY在系统发育树中关系较近。
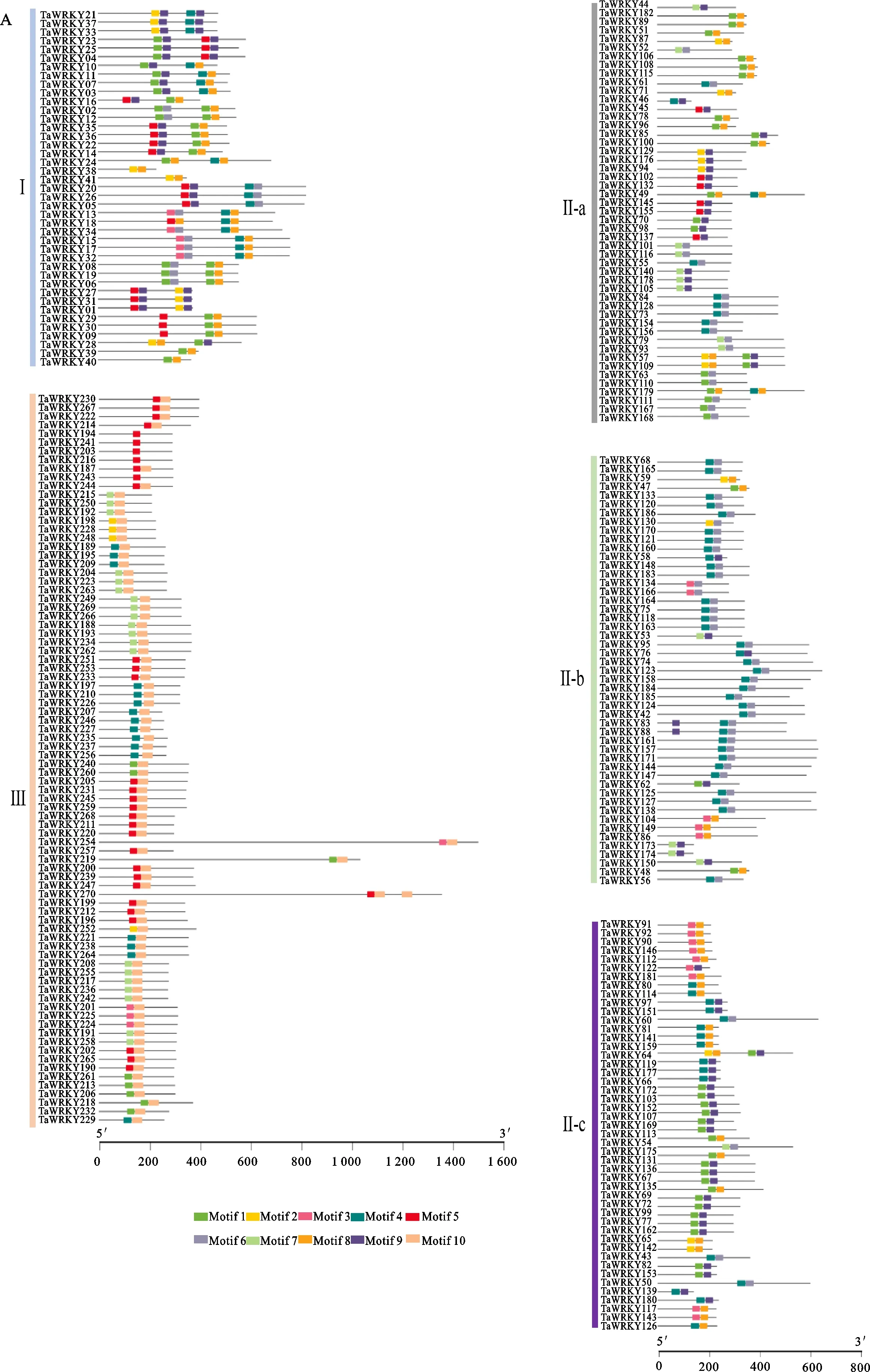
图2 TaWRKY蛋白的基序分析
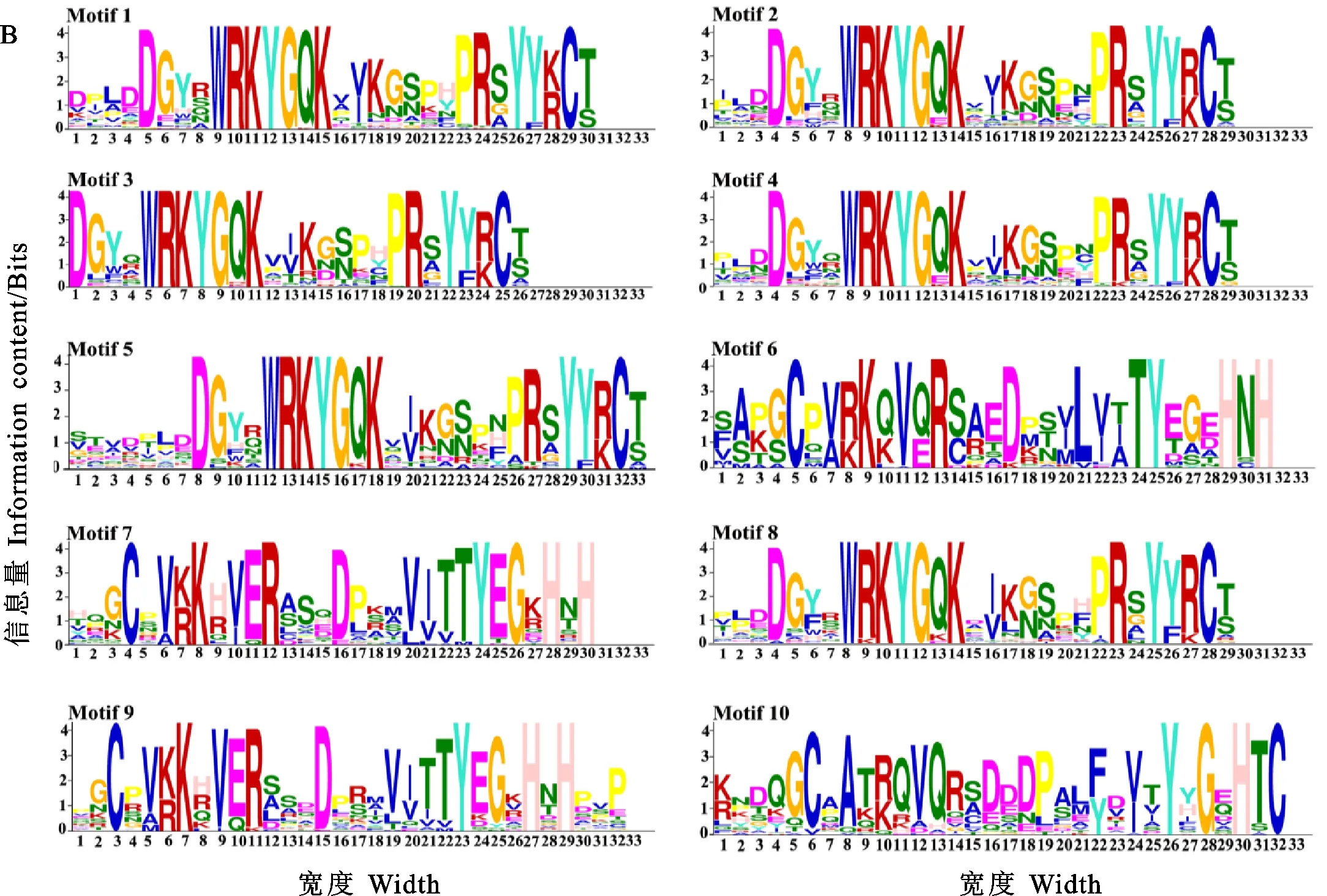
A:TaWRKY蛋白的基序分布; B:TaWRKY蛋白的基序序列。氨基酸字母的高度代表在该位点相应氨基酸的相对频率。
2.4 TaWRKY基因的结构分析
在Bio-Linux环境下,通过Perl程序进行分析,用CSDS 2.0软件绘制基因结构图,结果(图3)表明,大部分TaWRKY基因包含1~5个内含子(除TaWRKY13包含8个内含子)。在同一进化分支的TaWRKY基因表现出类似的基因结构,如TaWRKY167和TaWRKY168。但也有一些例外,如TaWRKY39和TaWRKY40虽然聚类为同一分支,但TaWRKY40仅有1个内含子,而TaWRKY39含有3个内含子。

图3 部分TaWRKY的基因结构
2.5 TaWRKY基因在小麦胚性愈伤组织形成过程中的表达模式
用周麦18幼胚培养3、5和15 d获得的胚性愈伤组织(IME3、IME6、IME15)和成熟胚培养3、5和15 d获得的非胚性愈伤组织(ME3、ME6、ME15)构建转录组测序文库,对不同文库的转录组数据进行分析。结果表明,共鉴定获得39个TaWRKY基因(表2),其中27个在基因组数据库中鉴定到且在愈伤组织中表达,12个在小麦基因组数据中没有鉴定到,仅在小麦愈伤组织发育中特异表达(配对比例为0)。TaWRKY基因在不同转录组文库中的差异表达分析表明,共有11个TaWRKY基因差异表达,其中包含5个(TaWRKY275、TaWRKY276、TaWRKY277、TaWRKY280和TaWRKY281)特异表达,结果见表3。随机选择8个差异表达的TaWRKY基因(含3个特异表达)进行qRT-PCR验证,结果与转录组测序结果一致(表4)。
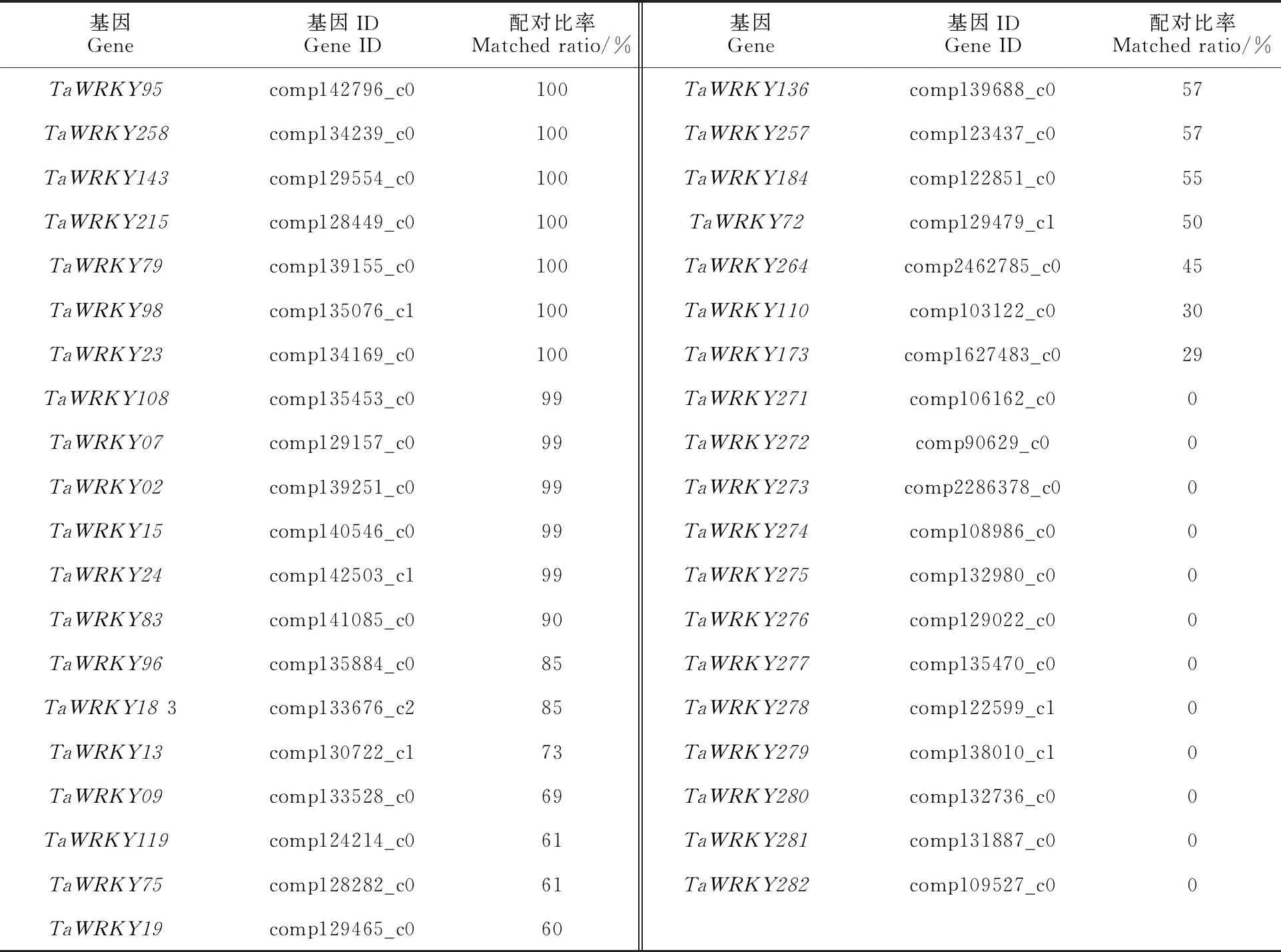
表2 小麦胚性愈伤组织形成中鉴定到的TaWRKY基因
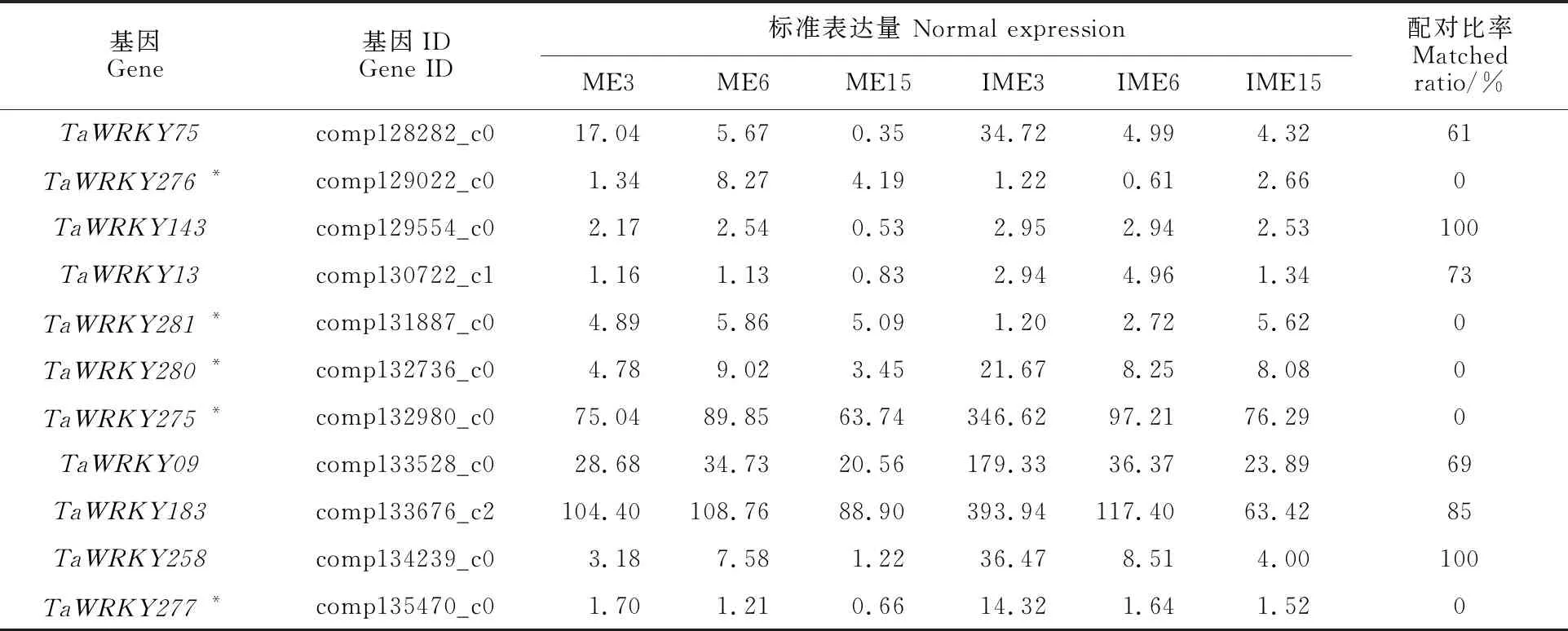
表3 小麦胚性愈伤组织形成中差异表达的TaWRKY基因

表4 随机选取的8个TaWRKY差异表达基因在小麦愈伤组织中的相对表达量
3 讨 论
WRKY转录因子在高等植物中广泛存在,家族成员较多,参与调控植物的多种生物学功能。随着植物基因组测序的逐渐完成,多种植物在全基因组范围内已经进行了WRKY转录因子家族成员的鉴定[13-19],而小麦WRKY转录因子的研究多以转录组或EST序列为基础[20-22],小麦基因组信息的公布为在全基因范围内进行小麦WRKY转录因子的研究提供了条件。本研究利用小麦基因组数据库,同时结合小麦胚性愈伤组织形成中的转录组数据,共鉴定获得270个小麦WRKY转录因子,根据WRKY结构域特征可分为Ⅰ、Ⅱ、Ⅲ三组[10],分别含有41、145和84个小麦WRKY转录因子,均多于拟南芥、水稻、棉花等植物中鉴定到的WRKY转录因子数量[11-12,31-32],可能这与小麦是六倍体及其具有庞大的基因组有关。
在研究方法上,本研究用Hummer搜索和NCBI Blast两种方法相结合来鉴定小麦WRKY转录因子,可能也是鉴定到小麦WRKY数量较多的原因之一。
在小麦胚性愈伤组织发育过程中,共鉴定到39个TaWRKY基因,其中12个在小麦愈伤组织发育过程中特异表达。在小麦胚性愈伤组织中鉴定到的小麦WRKY转录因子数量(39)远小于小麦全基因组条件下鉴定到的小麦WRKY转录因子数量(270),这可能与该数据是来源于特定发育时期的材料以及转录组测序的程度有关。
WRKY结构域中存在高度保守的WRKYGQK序列,但也存在变异,如在水稻中,WRKYGEK和WRKYGKK是WRKY转录因子中WRKY结构域两个常见的变异[31,33]。本研究在小麦中也发现了WRKYGEK和WRKYGKK两种变异,其中有13个小麦WRKY转录因子存在WRKYGEK变异(均位于第III组),有15个存在WRKYGKK变异(有3个位于第I组,该组中有两个WRKY结构域,一个变异,另一个不变异;有12个位于第II组)。
小麦胚性愈伤组织的形成是一个复杂的过程,本研究发现,在小麦胚性愈伤组织不同发育时期,有11个TaWRKY基因差异表达,其中5个特异表达,说明小麦WRKY转录因子与胚胎愈伤组织的形成相关,小麦WRKY转录因子通过何种途径对植物胚胎愈伤组织的形成进行调控还需进一步研究。
