PEI修饰的氢氧化铝纳米颗粒增强树突状细胞抗原提呈作用的初步研究
2021-11-05任宏艳康美云王子妤张璐瑶王永韧
任宏艳,康美云,王子妤,姚 琦,张璐瑶,王永韧*
1南京中医药大学医学院整合医学学院病理学与病理生理学系,江苏 南京 210023;2南京医科大学附属儿童医院血液肿瘤科,江苏 南京 210008
树突状细胞(dendritic cell,DC)疫苗因其在肿瘤免疫治疗中的应用而引起广泛关注。DC 是启动特异性免疫应答最强大的专职抗原提呈细胞(anti⁃gen presenting cell,APC)。然而,DC疫苗的临床疗效往往由于缺乏能够引起充分免疫应答的有效抗原而受到限制[1-2]。因此,增强DC对抗原的加工提呈能力是促进DC疫苗成功应用于临床的关键策略。
免疫佐剂常用来与抗原结合以增强其免疫原性进而诱导有效的免疫应答[3-4]。氢氧化铝是近年来应用最广泛的佐剂之一。此外,由于其卓越的安全性,含铝佐剂已被美国食品和药物管理局批准用于临床[3-7]。然而,经典的含铝佐剂只能触发适度的体液免疫应答,而无法诱导有效的细胞免疫应答。氢氧化铝纳米颗粒由微晶纳米纤维簇组成,具有优异的蛋白质吸附性能[3,8]。许多研究表明,氢氧化铝纳米颗粒具有有效表面积大、生物相容性好、载抗原能力强等显著优势[3,7-8]。我们之前的研究表明[9],当抗原共价结合到α⁃Al2O3纳米颗粒时,DC对抗原的交叉提呈效率显著提高。最近研究表明,聚乙烯亚胺(polyethylenimine,PEI)应用于基因传递系统时,可以产生较高的表面电荷和膜干涉效应,从而促进有效的基因转染[10-11]。本文通过PEI修饰的氢氧化铝纳米颗粒HS(HS/PEI)与抗原联合后负载DC以探索HS/PEI是否能增强DC对抗原的交叉提呈诱导特异性T细胞应答及其机制。
1 材料和方法
1.1 材料
实验动物选用6~8 周龄SPF 级C57BL/6 小鼠,雌性,体重18~20 g,购自扬州大学比较医学中心。6~8 周龄SPF 级OT⁃Ⅰ小鼠,雌性,体重18~20 g,购自南京大学模型动物研究中心。实验动物的饲养、使用和操作均获得南京中医药大学实验动物伦理委员会批准。细胞株及试剂mutuDC 细胞株和B3Z细胞株由美国波特兰肿瘤中心的胡红明教授馈赠。DMEM 培养基、RPMI⁃1640 培养基、胎牛血清、PBS(Gibco 公司,美国);卵清蛋白(ovalbumin,OVA)、PEI、CPRG(chlorophenol red⁃β⁃D⁃galactopy⁃ranoside)(Sigma 公司,美国);HS(Chemtrade Chemi⁃cals 公司,美国);蛋白定量检测试剂盒(Thermo Fisher Scientific 公司,美国);R848(Invivogen 公司,法国);白介素(interleukin,IL)⁃12检测试剂盒(eBio⁃science 公司,美国);OVA 抗体(Santa Cruz 公司,美国);His 标签蛋白纯化填料Ni⁃Sepharose excel(GE Healthcare 公司,美国);CFSE(carboxyfluorescein succinimidyl ester)(BioLegend公司,美国)。
1.2 方法
1.2.1 细胞培养及HS/PEI⁃OVA的制备
来源于CD11c:SV40LgT转基因C57BL/6小鼠脾脏的mutuDC细胞株在含10%胎牛血清、100 U/mL青霉素、100 mg/mL 链霉素、2 mmol/L L⁃谷氨酰胺的DMEM 培养基中培养,B3Z 细胞株(表达LacZ 的H⁃2Kb⁃OVA257⁃264特异性CD8+T细胞杂交瘤)在含10%胎牛血清、100 U/mL 青霉素、100 mg/mL 链霉素、2 mmol/L L⁃谷氨酰胺的RPMI⁃1640 培养基中培养。将0.5 mL HS(5 mg/mL)加入1.5 mL PEI(8 000 Da,25 mg/mL)后与OVA 溶液室温下共孵育1 h 制备成HS/PEI⁃OVA混合液。
1.2.2 CPRG法检测DC对抗原的提呈能力
mutuDC 细胞(2×104个,50 μL/孔)和刺激剂[干扰素(interferon,IFN)⁃α、R848]作用前后的HS/PEI⁃OVA(10 μg/mL,50 μL/孔)混合后放入96 孔U 底培养板,6 h后与B3Z细胞(2×105个,100 μL/孔)37 ℃共孵育过夜。离心,取上清,200 μL PBS清洗细胞后加入CPRG工作液,37 ℃孵育4 h后加入终止液(50 μL/孔),850g离心7 min后取上清,酶标仪595 nm波长检测吸光度。B3Z 细胞特异性识别OVA 激活其中LacZ的表达,与CPRG显色底物结合后生成氯酚红,通过测定吸光度检测B3Z细胞的活化情况。
1.2.3 Western blot检测DC中泛素化OVA蛋白的表达
mutuDC(5×106个)分别与PEI⁃OVA、HS⁃OVA、HS/PEI⁃OVA(10 μg/mL)于37 ℃共孵育6 h后以PFO(perfringolysin O,100 ng/mL)处理30 min,离心后收集上清,蛋白定量检测试剂盒测定上清中的蛋白总量。按课题组之前的方法富集PFO提取液中的泛素化蛋白[12]。将带有His6标记的泛素化蛋白纯化工具Vx3GFP融合蛋白(30 μg/mL)加入等蛋白总量的PFO提取液中,4 ℃孵育过夜。加入Ni⁃Sepharose excel层析柱颗粒,4 ℃旋转混合1 h后将混合物移至层析管中进行亲和层析纯化。离心后,用含5 mmol/L 咪唑的Tris⁃NaCl 缓 冲液(20 mmol/L Tris⁃Cl,300 mmol/L NaCl,pH 8.0)清洗树脂,最后用含250 mmol/L 咪唑的Tris⁃NaCl缓冲液洗脱his6⁃Vx3GFP结合的泛素化蛋白。洗脱液在4 ℃下以PBS 透析过夜,将富集的泛素化蛋白保存在-80 ℃。通过Western blot检测其中泛素化OVA 蛋白的含量。将富集的泛素化蛋白经SDS⁃PAGE分离,转移到PVDF膜,加入OVA抗体4 ℃孵育过夜,TBST 洗膜3 次后加入HRP 标记的二抗室温下孵育1 h,TBST 洗膜3 次后加化学发光试剂,在暗室中采用凝胶成像分析仪进行分析。
1.2.4 ELISA法检测DC负载HS/PEI⁃OVA后培养上清中IL⁃12的表达
mutuDC 细胞(2×104个,50 μL/孔)和刺激剂(IFN⁃α、R848)作用前后的HS/PEI⁃OVA(10 μg/mL,50 μL/孔)混合后放入96孔U底培养板37 ℃孵育6 h后,收获细胞培养上清。ELISA 法检测培养上清中IL⁃12的表达。
1.2.5 CFSE标记的OT⁃ⅠT细胞增殖实验
取OT⁃Ⅰ小鼠脾脏细胞加入CFSE染料(终浓度10 μmol/L),37 ℃孵育10 min,加入5 倍体积的预冷PBS,冰上放置5 min,低温离心10 min 后以RPMI⁃1640培养液重悬至4×106个/mL;2×105个DC分别与PEI⁃OVA、HS⁃OVA、HS/PEI⁃OVA(10 μg/mL)于37 ℃共孵育6 h后与上述CFSE标记好的OT⁃Ⅰ小鼠脾脏细胞(4×106个)混合37 ℃共孵育5 d,流式细胞术检测CFSE标记的OT⁃ⅠT细胞的增殖。
1.2.6 HS/PEI⁃OVA 免疫后小鼠体内CD8+IFN⁃γ+T细胞比率及特异性T细胞分泌IFN⁃γ的检测
C57BL/6小鼠左下侧腹部下注射接种B16⁃OVA细胞(2×105个/100 μL)。待移植瘤生长的第7天,移植瘤有米粒大小时,随机分组,每组5只,两侧腹股沟淋巴结注射OVA、HS⁃OVA、HS/PEI⁃OVA(每侧15 μg/15 μL),以PBS 作为对照组;第12天尾静脉注射DC⁃OVA、DC⁃HS⁃OVA、DC⁃HS/PEI⁃OVA 疫苗(每只5×106个/500 μL),对照组注射DC⁃PBS(每只5×106个/500 μL)。第16天,取小鼠脾脏和淋巴结细胞,含10%胎牛血清的RPMI⁃1640重悬细胞后放入48孔板(每孔2×106个/mL),每孔加SIINFEKL(1 μg/mL,OVA257⁃264peptide)或CD3抗体(5 μg/mL)再刺激12 h后,每孔加50 ng/mL 佛波酯、500 ng/mL 钙离子载体和2 μmol/L 高尔基体抑制剂继续培养6 h,收获细胞,流式细胞术检测CD8+IFN⁃γ+T 细胞比率;刺激72 h后,收取培养上清,ELISA检测培养上清中IFN⁃γ的水平。CD3抗体作为阳性对照。
1.3 统计学方法
实验数据用GraphPad Prism 7.0 软件进行统计学分析,各组数据用均数±标准差()表示,多组定量资料比较采用单因素方差分析(one⁃way ANO⁃VA)检验,多组间数据两两比较采用LSD 法。两组定量数据比较用t检验。P<0.05 为差异有统计学意义。
2 结果
2.1 HS/PEI 显著增强mutuDC对OVA 蛋白的抗原提呈作用
为了验证HS/PEI 在DC 抗原提呈中的作用,将不同浓度(0.001、0.003、0.010、0.030、0.100、0.300、1.000、3.000、10.000、30.000、100.000、300.00 μg/mL)的PEI、HS、HS/PEI 分别与OVA 蛋白(10 μg/mL)室温共孵育1 h,mutuDC 负载PEI⁃OVA、HS⁃OVA、HS/PEI⁃OVA 后与B3Z 细胞37 ℃孵育过夜,CPRG 法检测DC对抗原的提呈能力。结果发现,随着纳米颗粒浓度上升,与PEI⁃OVA、HS⁃OVA组相比,HS/PEI⁃OVA组的吸光值显著上升(图1A、B)。并且当纳米颗粒浓度为3 μg/mL 时,mutuDC 负载HS/PEI⁃OVA对B3Z细胞的激活已显著高于其他组(图1B),故在后续实验中,OVA蛋白浓度皆为10 μg/mL,纳米颗粒浓度皆为3 μg/mL。提示HS/PEI 可显著增强mutuDC对OVA抗原的提呈能力。
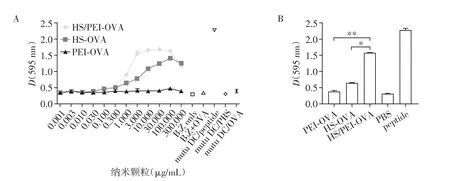
图1 CPRG法检测DC分别负载PEI⁃OVA、HS⁃OVA、HS/PEI⁃OVA后对OVA的提呈能力Figure 1 The efficiency of OVA cross⁃presentation by DCs loaded with PEI⁃OVA,HS⁃OVA or HS/PEI⁃OVA was deter⁃mined by CPRG assay
为了探讨HS/PEI 增强DC 抗原提呈作用的机制,mutuDC 与PEI⁃OVA、HS⁃OVA、HS/PEI⁃OVA 于37 ℃共孵育6 h 后加入PFO 处理DC。课题组以往研究已证实泛素化蛋白能增强DC的交叉提呈刺激特异性T细胞活化[12]。因此我们运用泛素化蛋白纯化工具Vx3GFP 融合蛋白从上述PFO 处理的DC 上清液中富集泛素化蛋白,Western blot检测其中泛素化OVA 蛋白的表达。结果显示,DC 负载HS/PEI⁃OVA 后泛素化OVA 蛋白的表达明显高于PEI⁃OVA和HS⁃OVA组(图2A)。
成熟的DC 分泌多种细胞因子,其中IL⁃12可通过促进交叉提呈诱导有效的T细胞应答[13-14]。为进一步探讨HS/PEI⁃OVA刺激对DC分泌IL⁃12的影响,将mutuDC 分别与PEI⁃OVA、HS⁃OVA、HS/PEI⁃OVA于37 ℃共孵育6 h后收取上清,ELISA检测细胞培养上清中IL⁃12的表达。结果显示,HS/PEI⁃OVA 刺激DC分泌IL⁃12的水平明显高于其他两组(图2B)。

图2 HS/PEI增强DC抗原提呈能力及IL⁃12的分泌Figure 2 Enhanced cross⁃presentation and secretion of IL⁃12 in DCs by HS/PEI
以上结果提示PEI修饰的经典纳米铝佐剂可显著增强DC对抗原的交叉提呈,诱导有效的T细胞应答。
2.2 IFN⁃α和TLR 刺激剂R848对DC 提呈HS/PEI⁃OVA的影响
TLR 和I⁃IFN 可调节DC 的成熟、抗原提呈及其诱导的抗病毒和抗肿瘤反应[15-17]。为检测TLR和I⁃IFN对DC提呈HS/PEI⁃OVA的影响,mutuDC负载刺激剂(IFN⁃α、R848)作用前后的HS/PEI⁃OVA 于37 ℃孵育6 h后收取上清,ELISA检测细胞培养上清中IL⁃12的表达;与B3Z细胞37 ℃孵育过夜后,CPRG法检测DC对HS/PEI⁃OVA 的提呈能力。结果显示,与未刺激的HS/PEI⁃OVA组相比,IFN⁃α和R848 刺激组DC分泌IL⁃12的水平显著升高(图3A),且能进一步刺激B3Z 细胞的活化(图3B)。提示IFN⁃α、R848可进一步增强DC对HS/PEI⁃OVA的交叉提呈,诱导高效的T细胞应答。

图3 IFN⁃α和TLR 刺激剂R848对DC 提呈HS/PEI⁃OVA的影响Figure 3 Cross⁃presentation of HS/PEI⁃OVA by DCs can be enhanced by IFN⁃α and TLR agonist R848
2.3 DC负载HS/PEI⁃OVA后显著促进OT⁃ⅠT细胞的增殖
DC 将抗原加工处理提呈给T 细胞在刺激T 细胞活化的同时也会促进T 细胞的增殖。OT⁃Ⅰ转基因小鼠CD8+T细胞的T细胞受体(TCR)特异性识别H⁃2Kb⁃OVA257-264。将DC 负载PEI⁃OVA、HS⁃OVA、HS/PEI⁃OVA后与CFSE标记的OT⁃Ⅰ小鼠脾脏细胞37 ℃共孵育5 d,流式细胞术检测OT⁃ⅠT细胞的增殖。结果显示,DC 负载HS/PEI⁃OVA 诱导OT⁃ⅠT细胞的增殖(80.1%)明显高于PEI⁃OVA组(27.0%)和HS⁃OVA组(43.3%)(图4)。结果进一步证实HS/PEI 能显著增强DC 交叉提呈抗原诱导特异性T 细胞免疫应答。
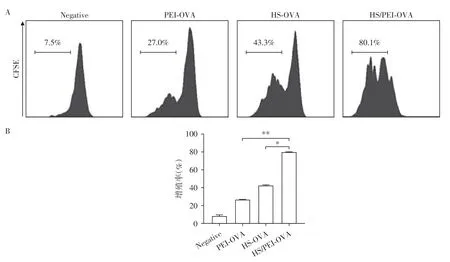
图4 DC负载HS/PEI⁃OVA后显著促进OT⁃ⅠT细胞的增殖Figure 4 The proliferation of OT⁃I T cells induced by DCs loaded with HS/PEI⁃OVA
2.4 DC⁃HS/PEI⁃OVA 疫苗显著增强小鼠体内CD8+IFN⁃γ+T细胞比率及特异性T细胞IFN⁃γ的分泌
为了观察DC⁃HS/PEI⁃OVA疫苗诱导小鼠体内T细胞活化的情况,我们建立小鼠B16⁃OVA黑色素瘤模型,第7 天分别给予OVA、HS⁃OVA 或HS/PEI⁃OVA 淋巴结初次免疫,第12 天DC⁃OVA、DC⁃HS⁃OVA 或DC⁃HS/PEI⁃OVA 尾静脉加强免疫(图5A)。第16天,取小鼠脾脏和淋巴结细胞,用SIINFEKL或CD3抗体再刺激12 h后,流式细胞术检测CD8+IFN⁃γ+T 细胞比率。结果显示,HS/PEI⁃OVA 免疫组的淋巴细胞在体外通过SIINFEKL 刺激后CD8+IFN⁃γ+T细胞比率明显高于OVA组、HS⁃OVA组和PBS对照组(图5B)。72 h 后,ELISA 检测细胞培养上清中IFN⁃γ水平。结果显示,与PBS 和OVA组相比,HS⁃OVA免疫组的IFN⁃γ水平有一定程度的升高;而HS/PEI⁃OVA免疫组IFN⁃γ水平增高更显著(图5C)。以上结果提示HS/PEI在体内可显著增强DC对抗原的交叉提呈诱导有效的T细胞应答。
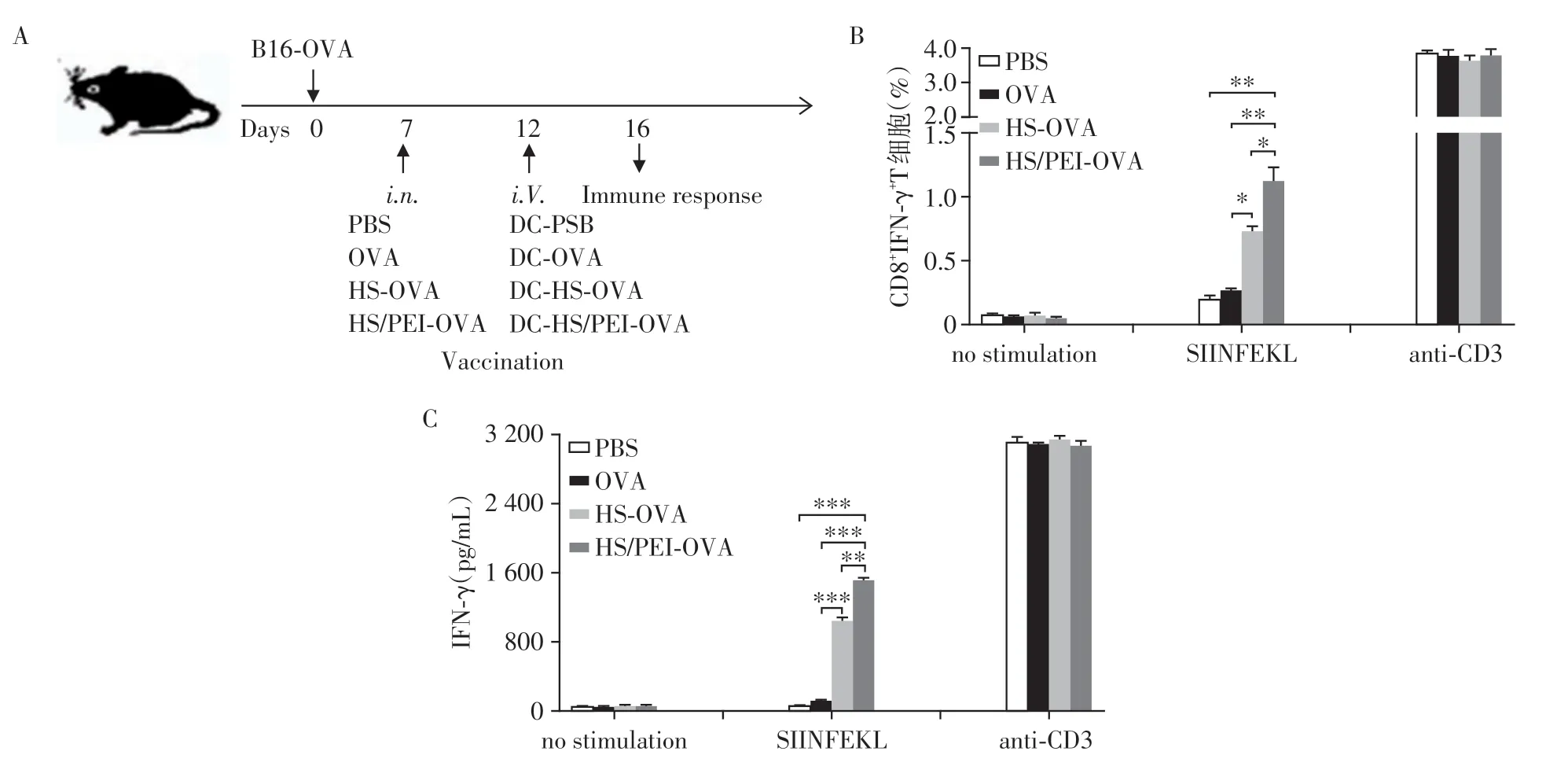
图5 DC⁃HS/PEI⁃OVA免疫B16⁃OVA荷瘤小鼠后显著增强小鼠体内CD8+IFN⁃γ+T细胞比率及特异性T细胞IFN⁃γ的分泌Figure 5 Vaccination with DC⁃HS/PEI⁃OVA significantly increased the percentage of CD8+IFN⁃γ+T cells and IFN⁃γ secre⁃tion of specific T cells in the B16⁃OVA tumor⁃bearing mice
3 讨论
一直以来铝佐剂作为免疫增强剂被广泛应用于疫苗[3-7]。经典的含铝佐剂只能增强抗体反应,而不能诱导有效的细胞免疫应答[18]。已有研究证实,氢氧化铝纳米颗粒具有很强的吸附能力,可以携带可溶性蛋白[3,8]。PEI由于携带强正电荷在修饰纳米颗粒时可增强其表面电荷[10]。本研究以OVA 作为模型抗原,以OVA 特异性CD8+T 细胞杂交瘤B3Z 为效应细胞,CPRG法检测B3Z细胞的活化。我们发现低浓度的OVA(10 μg/mL)与HS/PEI 混合后即可以引起较强的B3Z反应,提示PEI修饰的常规铝佐剂颗粒可以显著提高DC抗原交叉提呈的效率。
DC 中高浓度的抗原蛋白是诱导有效免疫应答的关键。本课题组已经证实,与细胞裂解液相比,泛素化蛋白能诱导更高效抗原交叉提呈[12]。Sims等[19]报道了一种可以结合泛素化蛋白的工具蛋白Vx3,本课题组前期构建的Vx3GFP工具蛋白已成功分离肿瘤细胞中的泛素化蛋白[12]。膜孔形成蛋白PFO 可以穿透细胞膜并释放胞浆蛋白到培养基中[20]。因此,本研究通过Western blot 检测了PFO 处理的DC胞浆中Vx3GFP 分离的泛素化抗原蛋白的表达情况。结果发现当OVA 与HS/PEI 结合后,DC 细胞内释放的Ub⁃OVA 明显增加。成熟DC 分泌的Th1 型细胞因子IL⁃12 已证实可以促进细胞毒性T 淋巴细胞(CTL)应答[13-14]。通过检测DC 负载OVA 联合纳米颗粒后分泌IL⁃12 的变化,我们发现DC 负载HS/PEI⁃OVA后分泌IL⁃12明显增加。表明HS/PEI可通过增加DC 中泛素化蛋白的表达及IL⁃12 的分泌诱导高效的细胞免疫应答。
已证实IFN⁃α/β能有效诱导DC的成熟,增强DC对抗原的交叉提呈从而激活CD8+T 细胞[16-17]。DC表达多种TLR,TLR的激活是DC实现有效交叉提呈的关键信号[21]。TLR7/8 激活剂R848、TLR7 激动剂Imiquimod刺激DC可显著上调DC表面MHC Ⅱ类分子和共刺激分子CD40、CD80、CD86的表达,并增加IFN⁃α的分泌[22-25]。本研究结果也证实了IFN⁃α和R848 刺激可进一步促进HS/PEI 增强DC 交叉提呈的作用。其他TLR刺激物Poly(I:C)、CpG、LPS等对HS/PEI增强DC交叉提呈的作用还有待进一步研究。
为了进一步探讨HS/PEI 能否在小鼠体内增强DC对OVA抗原的交叉提呈进而诱导特异性CD8+T细胞活化,我们用OVA、HS⁃OVA 或HS/PEI⁃OVA 负载DC 后免疫B16⁃OVA 荷瘤小鼠,取出小鼠脾脏和淋巴结细胞后用SIINFEKL 再刺激,发现HS/PEI⁃OVA 免疫组CD8+IFN⁃γ+T 细胞的比率明显增加,比其他组产生更高水平的IFN⁃γ。这些结果证明了HS/PEI在体内能显著增强DC交叉提呈抗原进而诱导特异性T细胞免疫应答。
抗原的高效提呈是诱导有效T细胞免疫应答的关键。本研究证明了PEI修饰的氢氧化铝纳米颗粒(HS/PEI)能显著增强DC 抗原提呈作用诱导高效的T 细胞应答。更重要的是,我们进一步证明了HS/PEI联合抗原后能够在体内诱导抗原特异性的免疫应答。DC负载HS/PEI联合抗原后在抗肿瘤免疫应答方面的作用还需进一步研究。
