楮果总黄酮体外抗氧化及对CCl4致脑损伤小鼠的保护作用
2021-10-29高志丽王瑞雯崔嘉纹白亚楠史海龙
高志丽,王瑞雯,崔嘉纹,白亚楠,史海龙,李 洋
(华北理工大学药学院,河北唐山 063000)
许多污染物如药物、杀虫剂、烟草烟雾、有机溶剂和有毒物质等都会引起氧化应激[1],使体内活性氧自由基的生成和细胞抗氧化防御体系之间失衡,危害机体健康。由于大脑代谢率很高且主要为有氧代谢,会产生大量活性氧自由基,更容易发生氧化应激[2],对脑组织造成氧化损伤。多种脑部疾病如心脑血管疾病、阿尔兹海默症等的发生发展机制,都涉及到氧化应激损伤[3]。抗氧化剂能抑制自由基的生成,改善机体抗氧化防御体系,延缓慢性疾病的发展[4]。因此,研究安全、有效、无毒、无副作用、能预防或减轻氧化应激的天然抗氧化剂具有重要意义。国内外研究表明,黄酮类化合物在体内外具有清除活性氧自由基、抑制脂质过氧化、增强体内抗氧化的能力[5],是天然抗氧化剂研究的热点。
楮果是桑科构属植物构树(Broussonetia papyriferaL.)的成熟果实,无毒、可食用,在我国分布非常广泛,资源丰富[6],在民间已有上千年的历史,是我国传统民间草药和保健食品。楮果中含有矿物元素、多糖、维生素、脂肪油、生物碱、色素等多种化学成分[7],其中色素呈橙红色,色泽鲜艳,稳定性较好,经初步分析属于黄酮醇类黄酮化合物[8]。楮果在改善记忆[9]、保护神经元[10]、抗氧化[11]等方面表现出良好的活性,但目前国内外对楮果总黄酮的研究还集中于提取、含量测定等方面,关于其抗氧化活性、防治氧化应激的研究报道较少,其食用和药用价值尚未得到开发利用。
动物四氯化碳(carbon tetrachloride,CCl4)中毒是一种模拟氧化应激的实验模型,在肝脏、肾脏、大脑和血液等组织中都能引起自由基的产生,造成氧化应激损伤[12−13]。本研究将楮果总黄酮的体外抗脂质过氧化实验与CCl4诱导小鼠脑组织氧化损伤实验相结合,评价其体内外抗氧化能力,为楮果总黄酮在功能性食品、抗氧化保健品和化妆品方面的开发利用提供参考。
1 材料与方法
1.1 材料与仪器
SPF 级昆明雄性小鼠60 只 体重(20±2)g,购自北京华阜康生物科技股份有限公司,许可证号:SCXK(京)2019-0008,饲养于华北理工大学实验动物中心,饲养环境12 h 黑暗的光照周期,温度在21~23 ℃和湿度在50%~60%,给予喂食认证的饮食和纯净水;楮果 采摘于唐山,经华北理工大学药学院刘占军教授鉴定为桑科构树属植物构树(Broussonetia papyriferaL.)的果实;肝素钠 美国biosharp 公司;过氧化氢(hydrogen peroxide,H2O2) 天津凯通化学试剂有限公司;L-抗坏血酸、硫代巴比妥酸 国药集团化学试剂有限公司;三氯乙酸 北京迈瑞达科技有限公司;芦丁(纯度>95%) 中国药品生物制品检定所;水飞蓟素 美国Sigma 公司;CCl4分析纯,天津市北方天医化学试剂厂;丙二醛(malondialdehyde,MDA)、谷胱甘肽(glutathione,GSH)、超氧化物歧化酶(superoxide dismutase,SOD)、谷胱甘肽过氧化物酶(glutathione peroxidase,GSH-PX)、过氧化氢酶(catalase,CAT)试剂盒 南京建成生物工程研究所;苏木素-伊红染色液 珠海贝索生物技术有限公司。
Lambda 35 紫外可见分光光度计 美国Perkin-Elmer 公司;VCX 130 超声匀浆器 美国Sonics &Materials 公司;ST 16R 冷冻离心机 美国Thermo Fisher 公司;ELX800 酶标仪 美国伯腾仪器有限公司;BX53F 显微镜 日本OLYMPUS 公司。
1.2 实验方法
1.2.1 楮果总黄酮的制备与含量测定 楮果洗净晾干,加入80%乙醇(料液比1:10 g/mL)在40 ℃、640 W超声下提取30 min,反复提取3 次,合并滤液,减压浓缩,得楮果总黄酮粗提液[14];称取活化的D-101 大孔树脂3 g(湿重),加楮果总黄酮粗提液50 mL,置于转速140 r/min、25 ℃恒温振荡器上,振荡吸附12 h后,用去离子水清洗吸附了黄酮的D-101 大孔树脂,然后加入50 mL 80%乙醇溶液,置于恒温振荡器上解吸24 h,抽滤,保存滤液(解吸液)[15]。解吸液减压浓缩后冻干,得楮果总黄酮粉末。参照《中国药典》2015 年版第一部“山楂叶”项下总黄酮的含量测定方法,以芦丁标准品为对照,用NaNO2−Al(NO3)3−NaOH 分光光度法绘制回归曲线,得到回归方程为:y=11.152x+0.0001,相关系数R2=0.999。按以上方法测得楮果总黄酮纯度达63.6%。
1.2.2 楮果总黄酮体外抗氧化能力的测定
1.2.2.1 对红细胞溶血抑制率的测定 昆明种小鼠禁食过夜,摘眼球采血,加入肝素钠抗凝。血样中加10 倍磷酸盐缓冲液(pH7.4),2000 r/min 离心5 min,沉淀物按上述方法离心洗涤3 次,制备为0.5%的红细胞悬液,于4 ℃保存(制备6 h 内使用)。
取1 mL 红细胞悬液,加0.1 mL 不同浓度的楮果总黄酮溶液(对照组加去离子水0.1 mL)和0.1 mL H2O2(100 mmol/L),37 ℃恒温水浴1 h,加入4 mL磷酸盐缓冲液,2000 r/min 离心5 min,415 nm 测吸光度,以抗坏血酸为阳性对照[16]。红细胞溶血抑制率计算公式如下:

式中:A415(对照)为空白组吸光度;A415(样品)为样品组吸光度。
1.2.2.2 对肝、脑组织匀浆的脂质过氧化抑制率的测定 取新鲜小鼠肝、脑组织,用预冷磷酸盐缓冲液冲洗血渍,擦干,称重。按料液比1:9的比例加磷酸盐缓冲液,将肝、脑组织于冰上剪碎、研磨,制成10%组织匀浆,3500 r/min 离心15 min,上清液4 ℃保存备用(用时稀释)。
取10%肝匀浆和2%脑匀浆各1 mL,分别加入0.1 mL 不同浓度楮果总黄酮溶液(对照组加去离子水),混匀,37 ℃水浴30 min,然后加0.1 mL H2O2(100 mmol/L),再水浴30 min,最后加入20%三氯乙酸和0.67%硫代巴比妥酸各1 mL,混匀后置于95 ℃水浴15 min,冷却后以5000 r/min 离心10 min,在532 nm 处测定上清液吸光度,用抗坏血酸作为阳性对照[17]。脂质过氧化抑制率计算公式如下:

式中:A532(对照)空白组吸光度;A532(样品)样品组吸光度。
1.2.2.3 线粒体肿胀度的测定 在4 ℃条件下,将10%肝匀浆1000 r/min 离心20 min,再将上清液10000 r/min 离心20 min,所得沉淀再加10 倍体积磷酸盐缓冲液10000 r/min 离心20 min,最后用冰磷酸盐缓冲液重悬制成悬液[18]。考马斯亮蓝染色法测定其蛋白浓度,调整为0.5 mg/mL 线粒体悬液,4 ℃保存。
取线粒体悬液2 mL 加不同浓度的楮果总黄酮溶液0.2 mL,再加FeSO4溶液(0.5 mmol/L)和抗坏血酸溶液(0.5 mmol/L)各0.2 mL,混匀后置于37 ℃恒温水浴锅,每隔5 min 在520 nm 测吸光度[18]。空白组用等体积磷酸盐缓冲液代替黄酮溶液、FeSO4溶液和抗坏血酸溶液,模型组用等体积磷酸盐缓冲液代替黄酮溶液,用芦丁作为阳性对照。
1.2.2.4 总抗氧化能力的测定 依次向试管中加H2SO4(3.0 mol/L)、Na3PO4(0.14 mol/L)和钼酸铵(0.02 mol/L)各1.0 mL,然后加入1.0 mL 不同浓度的楮果总黄酮溶液(空白组加等体积去离子水),再加入1.0 mL去离子水,混匀后95 ℃水浴加热90 min,695 nm 测吸光度[19]。用抗坏血酸作为阳性对照。
1.2.3 楮果总黄酮对CCl4致小鼠脑损伤的防护作用
1.2.3.1 小鼠脑损伤模型的建立 将60 只昆明种小鼠随机分为6 组(每组10 只):空白组、模型组、阳性对照组(水飞蓟素,0.2 g/kg)、楮果总黄酮低剂量组(0.15 g/kg)、楮果总黄酮中剂量组(0.3 g/kg)、楮果总黄酮高剂量组(0.6 g/kg),灌胃给药(楮果总黄酮低、中、高剂量组灌胃剂量设置依据预实验结果),1次/d,空白组和模型组每天灌胃10 mL/kg 等体积羧甲基纤维素(CMC)。除空白组外,其余各组隔天腹腔注射1:1 CCl4花生油溶液2 mL/kg,空白组给予同体积花生油,持续14 d[4,20]。末次给药后,禁食24 h处死小鼠,迅速取出脑组织。将右脑固定在4%多聚甲醛中以制备石蜡切片,将其余部分立即储存在−80 ℃用于其他测定。
1.2.3.2 测定氧化应激指标 取脑组织称重,按重量体积比(1:9)加入生理盐水,并于冰上剪碎,研磨制备为10%脑组织匀浆。3500 r/min 离心15 min,取适量上清液用于检测氧化应激指标MDA、GSH 含量及SOD、GSH-PX、CAT 活性,具体步骤按照相应试剂盒说明书进行。
1.2.3.3 组织病理学分析 取空白组、模型组、水飞蓟素组和楮果总黄酮各剂量组小鼠脑组织,固定于4%多聚甲醛中,石蜡包埋,切片,二甲苯、梯度乙醇脱蜡透明,HE 染色,中性树胶封固。光镜下观察脑组织病理变化。
1.3 数据处理
2 结果与分析
2.1 体外抗氧化能力
2.1.1 楮果总黄酮对红细胞的保护作用 红细胞具有很典型的生物体细胞膜结构,常用于分析和筛选天然抗氧化剂。在H2O2诱导下可以产生活性很强的·OH,诱发多不饱和脂肪酸氧化,破坏生物膜的结构和功能,从而造成红细胞溶血[16]。由图1 可见,在实验浓度0.1~3.2 mg/mL 之间,楮果总黄酮与抗坏血酸都表现出抑制小鼠红细胞溶血活性,且抑制活性随浓度的增加而增强。楮果总黄酮和抗坏血酸抑制红细胞溶血的IC50值分别为1.14 mg/mL 和0.43 mg/mL,虽然楮果总黄酮的IC50大于抗坏血酸,但当浓度为3.2 mg/mL 时,其对红细胞溶血的抑制作用与抗坏血酸已无明显差异,说明楮果总黄酮对红细胞溶血有较强的抑制作用。

图1 楮果总黄酮对红细胞溶血的抑制能力Fig.1 Inhibitory effect of total flavonoids from paper mulberry fruits on erythrocyte hemolysis
2.1.2 楮果总黄酮对肝、脑匀浆脂质过氧化抑制活性 脂质过氧化是多不饱和脂肪酸在细胞膜上的氧化过程,是氧自由基引起的主要有害变化之一[21]。不饱和脂肪酸的氧化分解会导致脂质过氧化链式反应,生成丙二醛等小分子产物容易引起细胞形态和功能的改变[18]。由图2 可见,在实验浓度范围(0.4~12.8 mg/mL)内,楮果总黄酮对肝脏脂质过氧化有一定的抑制作用,抑制率随浓度的增加而增加;其IC50值为1.19 mg/mL,抗坏血酸IC50值为1.06 mg/mL。当浓度大于6.4 mg/mL 时,楮果总黄酮对肝脂质过氧化的抑制率达90%以上,大于抗坏血酸对肝脂质过氧化的抑制率,说明楮果总黄酮对肝组织脂质过氧化有较强的抑制作用。

图2 楮果总黄酮对肝匀浆脂质过氧化的抑制能力Fig.2 Inhibitory effect of total flavonoids from paper mulberry fruits on lipid peroxidation of liver homogenate
由图3 可见,在实验浓度范围内,楮果总黄酮对脑匀浆脂质过氧化现象的抑制作用迅速增长,且呈现剂量依赖关系;其IC50值为0.87 mg/mL,抗坏血酸IC50值为7.88 mg/mL,说明楮果总黄酮对脑组织匀浆脂质过氧化的抑制作用远高于抗坏血酸。

图3 楮果总黄酮对脑匀浆脂质过氧化的抑制能力Fig.3 Inhibitory effect of total flavonoids from paper mulberry fruits on lipid peroxidation of brain homogenate
2.1.3 楮果总黄酮对线粒体肿胀度的抑制作用 在自由基存在下,线粒体膜极容易受到自由基的攻击,导致脂质过氧化,致使膜结构被破坏,线粒体通透性发生改变,从而引发线粒体肿胀[22]。由图4 可见,模型组的吸光度随着时间的增长呈明显下降趋势,楮果总黄酮2、4 mg/mL 组和芦丁实验组吸光度下降幅度明显降低,楮果总黄酮浓度越大吸光度下降幅度越小,同浓度下楮果总黄酮较阳性对照芦丁组的吸光度下降幅度小,表明楮果总黄酮对线粒体肿胀有抑制作用,且对线粒体的保护作用强于芦丁。

图4 楮果总黄酮对线粒体肿胀的抑制能力Fig.4 Inhibitory effect of total flavonoids from paper mulberry fruits on mitochondrial swelling
2.1.4 楮果总黄酮的总抗氧化力 用磷钼酸盐络合物法测样品的总抗氧化力,该方法的原理是钼在硫酸和磷酸的作用下发生还原反应,产物为绿色,在695 nm 处有较强的吸收峰[23]。而抗氧化剂会与钼竞争发生还原反应,其抗氧化的能力可由生成钼蓝的量间接反映,吸光度越大代表总抗氧化能力也越大。由图5 可知,随着楮果总黄酮浓度的增加,总抗氧化能力逐渐增强,抗氧化能力与浓度之间存在明显的量效关系,说明楮果总黄酮具有一定的抗氧化能力。

图5 楮果总黄酮的总抗氧化能力Fig.5 Total antioxidant capacity of total flavonoids from paper mulberry fruits
2.2 楮果总黄酮对CCl4 致脑氧化损伤小鼠的保护作用
2.2.1 楮果总黄酮对小鼠脑组织MDA 含量的影响MDA 是细胞内脂质过氧化自由基的分解产物,是脂质过氧化和氧化应激的重要生物标志物之一,产生过量会破坏生物膜的结构和功能[24]。MDA 含量可反映组织细胞脂质过氧化的速率或强度。如图6 所示,与空白组相比,模型组MDA 含量极显著增加(P<0.01),表明CCl4诱导的小鼠脑氧化损伤模型成功建立。与模型组相比,连续灌胃楮果总黄酮后,楮果总黄酮中、高剂量组小鼠脑组织MDA 含量极显著降低(P<0.01),降低程度与灌胃剂量成明显的量效关系,可见楮果总黄酮能降低小鼠脑组织脂质过氧化反应,防护脑组织氧化损伤。

图6 楮果总黄酮对小鼠脑组织MDA的影响(,n=10)Fig.6 Effects of total flavonoids from paper mulberry fruits on MDA content in brain tissue of mice (,n=10)
2.2.2 楮果总黄酮对小鼠脑组织GSH 含量的影响GSH 是一种重要的非酶促抗氧化剂,在细胞内和细胞外都能对抗外来生物和中和活性氧,因此,体内GSH 水平被认为是其抗氧化能力的重要指标[20,25]。如图7 所示,模型组小鼠脑组织GSH 含量较空白组显著降低(P<0.01),表明CCl4能破坏机体抗氧化防御机制。连续灌胃楮果总黄酮后,各剂量组小鼠脑组织GSH 含量显著提高(P<0.01),表明楮果总黄酮能显著增强小鼠脑组织的抗氧化能力。同时,楮果总黄酮高剂量组的GSH 含量与空白组相比已无显著性差异(P>0.05),可见高剂量的楮果总黄酮对组织抗氧化能力的恢复作用效果出色。

图7 楮果总黄酮对小鼠脑组织GSH 含量的影响(,n=10)Fig.7 Effect of total flavonoids from paper mulberry fruits on GSH content in brain tissue of mice (,n=10)
2.2.3 楮果总黄酮对小鼠脑组织中SOD、GSH-PX和CAT 活性的影响 SOD、GSH-PX 和CAT 等是体内重要的抗氧化酶,在清除活性氧和防止活性氧形成的过程中起着至关重要的作用。SOD 是机体抵抗活性氧的一种防御机制,在氧化与抗氧化平衡中起着重要的作用,是过氧阴离子自由基的清除剂,能抑制自由基诱导的脂质过氧化,保护细胞免受氧化应激损伤[26]。GSH-PX 部分位于细胞膜内,是抗脂质过氧化作用的酶保护系统之一[27−28]。GSH-PX 和CAT 分解H2O2,同时最大限度减少羟基自由基的潜在危害[2]。如图8 所示,模型组小鼠脑组织SOD、GSH-PX 和CAT 酶活性较空白组均明显降低,且差异具有统计学意义(P<0.01),表明给予小鼠CCl4后,机体出现氧化应激反应。与模型组相比,楮果总黄酮各组小鼠脑组织中SOD 和GSH-PX 活性均极显著升高(P<0.01),楮果总黄酮中、高剂量组小鼠脑组织CAT 活性显著升高(P<0.05 或P<0.01),且呈良好的剂量依赖关系,同时,楮果总黄酮高剂量组与空白组SOD 和GSHPX 活性相比已无显著差异(P>0.05),可见楮果总黄酮对CCl4诱导的小鼠脑组织氧化应激具有较明显的抑制作用;以上结果表明楮果总黄酮可通过提高CCl4致脑损伤小鼠脑组织中SOD、GSH-PX 和CAT 酶活性,发挥防护脑组织氧化应激损伤的作用,且剂量越高保护效果越好。

图8 楮果总黄酮对小鼠脑组织中SOD、GSH-PX 和CAT 活性的影响(,n=10)Fig.8 Effect of total flavonoids from paper mulberry fruits on SOD,GSH-PX,CAT activity in brain tissue of mice (,n=10)
2.2.4 病理组织学观察 如图9 可见,空白组脑组织海马区组织学表现正常,神经细胞数目多,排列紧密,均匀整齐,胞核全染;而模型组神经细胞结构疏松,数量减少,排列紊乱,核轮廓不规则,核异色、固缩、核仁消失、胞浆周围晕染、细胞间距大,说明脑组织发生了严重的氧化应激损伤。楮果总黄酮干预后,脑组织海马区神经细胞有不同程度的恢复。低剂量组海马区细胞疏松,部分神经细胞不饱满,神经细胞胞浆和细胞核界限不清;中剂量组海马区病理性形态学改变有明显改善;高剂量组海马区细胞较为致密,细胞数量和形态基本恢复正常。由此可见,楮果总黄酮对CCl4致小鼠脑氧化损伤具有显著的防护作用,防护作用与楮果总黄酮浓度呈明显的量效关系。
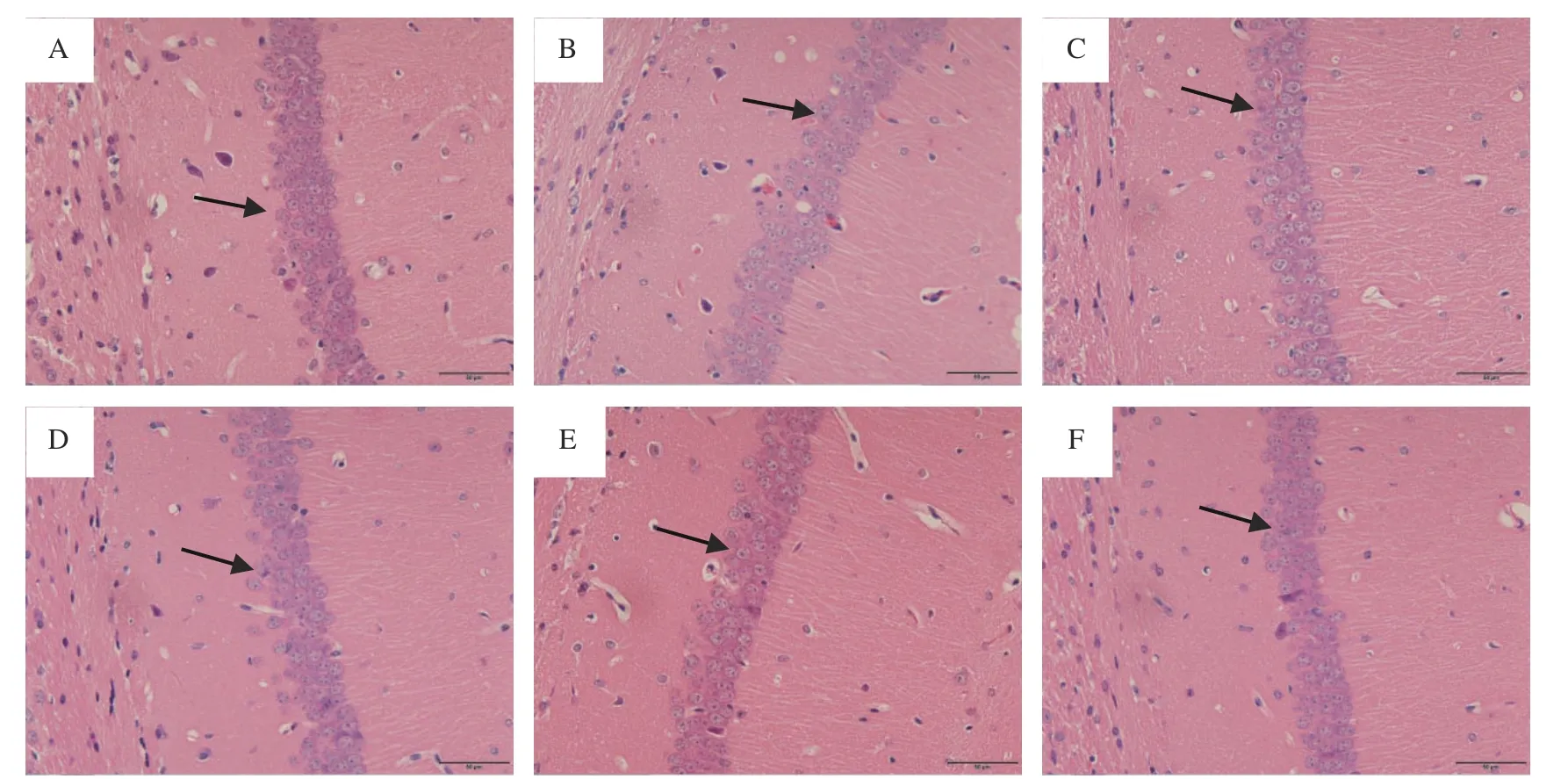
图9 楮果总黄酮对小鼠脑组织海马区病理形态学的影响(HE,400×)Fig.9 Effect of total flavonoids from paper mulberry fruits on pathomorphological of hippocampus in mice brain tissue (HE,400×)
3 结论
体外实验数据表明,楮果总黄酮对生物膜实验体系中红细胞溶血和肝组织脂质过氧化的抑制作用达到一定浓度时,大于阳性对照;而其对于脑组织脂质过氧化和线粒体肿胀的抑制作用均大于阳性对照,且抑制能力与浓度呈显著的量效关系,总抗氧化能力也很强。由此证明,楮果总黄酮具有较强的体外抗氧化能力。体内实验结果表明,低剂量楮果总黄酮对小鼠脑组织SOD、GSH 和GSH-PX 水平均有改善作用,对MDA 含量和CAT 活性无明显改善作用;而中、高剂量楮果总黄酮均能显著降低CCl4致氧化损伤小鼠脑组织中过氧化物质MDA 含量,升高抗氧化酶SOD、GSH-PX、CAT 活性,增加GSH 含量,提高机体防御能力,并修复脑组织海马区病理学变化。由此可见,楮果总黄酮对小鼠脑组织氧化损伤的保护作用是通过降低MDA 水平,升高GSH 含量,同时提升SOD、GSH-PX 和CAT 活性来实现的。有研究表明,黄酮类化合物槲皮苷通过降低MDA 水平并将SOD、GSH-PX 和CAT 酶的活性恢复到接近正常水平来发挥其对CCl4诱导小鼠脑损伤的保护作用[20]。这与本研究结果相一致。
综上所述,楮果总黄酮具有良好的体内外抗氧化活性,有助于保护机体免受氧化应激产生的有害影响,可作为新型天然抗氧化剂,对功能性食品、营养保健品方面的研究和开发具有巨大的经济价值。
