驴乳的抗氧化功能及作用机理
2021-08-09岳远西闫素梅
岳远西 闫素梅
(内蒙古农业大学动物科学学院,内蒙古自治区高校动物营养与饲料科学重点实验室,呼和浩特 010018)
随着人们生活水平的不断提高,消费者越来越意识到饮食与健康的关系,因而增加了其对食品营养价值的兴趣,对于含有功能成分的食品的需求也不断增加,这些功能成分在维持健康和预防疾病方面发挥着重要作用。驴乳中含有丰富的活性成分,包括溶菌酶、乳铁蛋白、乳过氧化物酶和免疫球蛋白等活性因子,具有抗氧化、抗肿瘤、抗菌及增强免疫力的作用[1-2]。早在1 300年前,我国的古代医学书籍中就记载了驴乳的药用价值,其味甘、冷利、无毒,可疗大热、止消渴[3]。Khan等[4]也指出,驴乳是具有抗氧化特性的功能性乳品,有益于人体的氧化防御,有可能在常规治疗中作为辅助佐剂,用于治疗心血管疾病、代谢紊乱、肠道健康等。有研究发现,与牛乳、绵羊乳和山羊乳相比,驴乳的氧化稳定性更高,具有较强的自由基清除能力和铁还原能力[5]。Lionetti等[6]研究表明,在大鼠饲粮中添加驴乳可增加其肝脏线粒体的解偶联功能,提高大鼠的抗氧化和抗炎防御能力。此外,有研究表明,与牛乳相比,驴乳具有更低的过敏性,更适应婴幼儿饮用[7-8],是理想的人乳代用品。而驴乳的抗氧化能力需要通过其包含的活性成分来实现。因此,本文综述了驴乳的基本成分组成、抗氧化功能及作用机制,为进一步研究驴乳的生理功能和开发利用提供理论参考。
1 驴乳的基本成分组成
驴乳含有乳蛋白、乳脂、乳糖、维生素和矿物元素等营养物质,其种类与牛乳等其他乳制品相似,其成分比例与之差异较大,但与人乳接近[9]。驴乳营养成分的最大特点是高乳糖、低乳脂,但不同资料报道的范围不尽一致。表1汇总了驴乳、马乳、牛乳和人乳的乳成分以及其乳蛋白组成。驴乳的乳糖含量为5.8%~7.4%,与人乳和马乳相似,高于牛乳。驴乳的乳脂含量为0.28%~2.00%,低于牛乳和人乳。在希腊、塞浦路斯地区的驴种以及中国德州驴的研究中,驴乳的乳脂含量仅为0.5%左右[10-11];而在Marchi等[12]的研究中,驴乳的乳脂含量甚至高达2.0%,这可能是由于地区、品种和饲粮的差异造成的。驴乳的乳蛋白含量为1.2%~2.0%,低于牛乳,但其蛋白质组成与牛乳差异很大。牛乳的乳蛋白中酪蛋白含量为80%,而驴乳中仅为30%~35%[13],而其乳清蛋白含量达60%以上,是名副其实的乳清蛋白乳。此外,驴乳中9种必需氨基酸的含量占氨基酸总量的42.52%,种类齐全,比例合理。驴乳中乳清蛋白部分含有6种主要蛋白质:乳铁蛋白(Lf)、血清白蛋白(SA)、β-乳球蛋白(β-Lg)、溶菌酶、α-乳蛋白(α-La)及免疫球蛋白Gs(IgGs)[14]。其中,溶菌酶含量远高于牛乳和人乳,是驴乳发挥特殊作用的主要活性物质之一[15]。驴乳中富含钙和硒[16-17],其中钙含量是人乳的2~3倍[18],钙磷比为1.7∶1.0,而硒含量是牛乳的5.16倍。驴乳含有大量的不饱和脂肪酸,人体必需脂肪酸亚油酸和亚麻酸的含量占脂肪酸总量的30.7%,分别比牛乳和人乳高出27和20个百分点。综上所述,驴乳的基本成分组成优于牛乳,与人乳最为接近,是理想的婴幼儿代乳品。
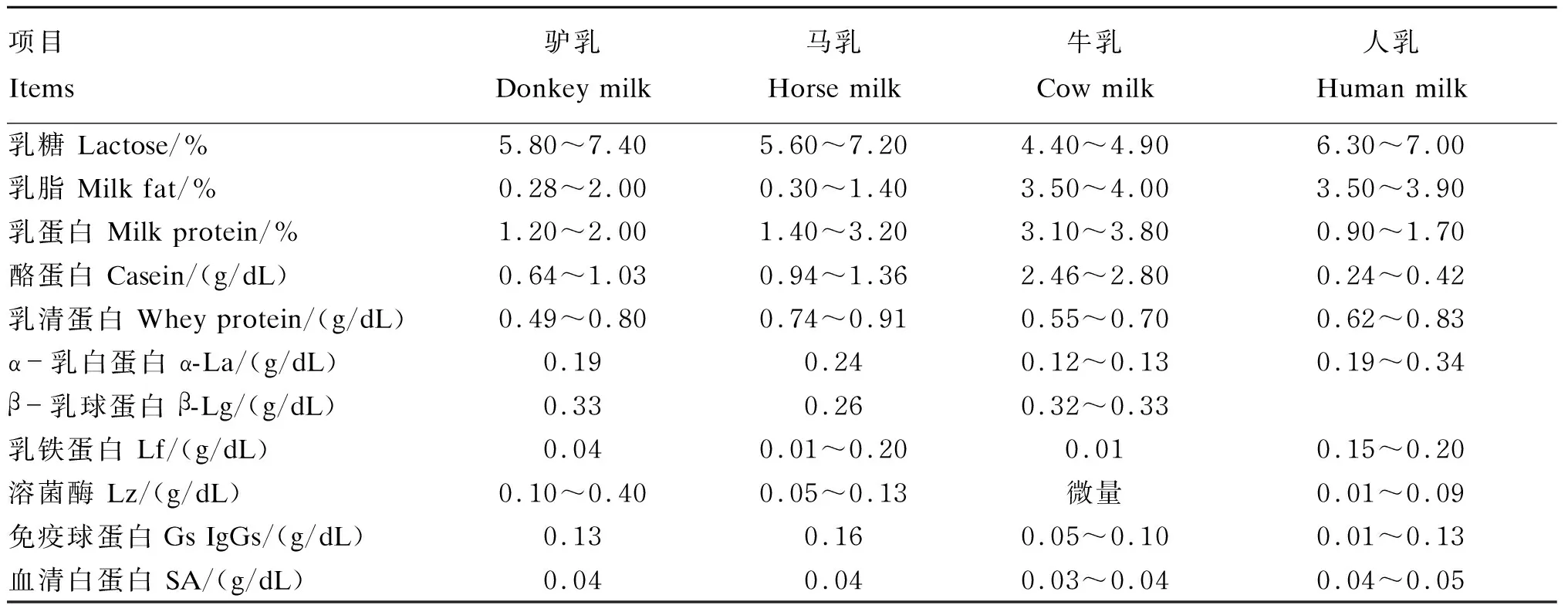
表1 驴乳、马乳、牛乳和人乳的乳成分比较
2 驴乳的抗氧化功能及作用机制
2.1 驴乳的抗氧化功能

2.2 驴乳的抗氧化作用机制
2.2.1 驴乳通过清除自由基发挥抗氧化功能

有研究发现,酪蛋白也具有抗氧化功能[25],其分子的一级结构对自由基发挥清除和淬灭作用[26]。Akan[27]研究了驴乳中酪蛋白的抗氧化作用,结果发现驴乳酪蛋白对·DPPH和ABTS+具有较强的清除能力。但目前关于驴乳酪蛋白抗氧化功能的研究报道很少见,具体结果需要进一步确定。
2.2.2 驴乳通过核因子E2相关因子2(nuclear factor erythroid E2 related factor 2,Nrf2)信号通路调节机体的抗氧化功能
Nrf2是一种重要的转录因子,在抗炎和抗氧化应激反应中发挥重要作用。Nrf2主要存在于细胞浆中,结合胞质接头蛋白Kelch样环氧氯丙烷相关蛋白1(Keap1)形成一种复合体。而Keap1是Nrf2的负性调控因子,其作用是介导Nrf2泛素化从而被蛋白酶降解,使Nrf2的生理活性维持在较低水平。当机体细胞遭受炎症或氧化刺激时,Nrf2磷酸化后可以与Keap1解离并活化,之后转移到细胞核内与相关抗氧化基因的启动子抗氧化反应元件(ARE)结合。这时,ARE受Nrf2诱导可调控下游靶基因,进而使GSH-Px和SOD合成增多,促进机体抗氧化能力的提升[32]。Trinchese等[2]对大鼠分别饲喂驴乳和人乳,结果发现大鼠机体内的过氧化氢含量减少,顺乌头酸酶活性受到抑制,而SOD活性增强,氧化还原状态[还原型谷胱甘肽(GSH)/氧化型谷胱甘肽(GSSG)比率]得到改善,研究认为这种结果可能归因于Nrf2-ARE通路的激活。Lionetti等[6]发现了类似的结果,饲喂驴乳提高了大鼠肝脏GSH含量,其线粒体中NQO1和GST活性增强。在轻度氧化应激下,Nrf2从其抑制剂Keap1中释放,可触发NQO1和GST编码基因的激活[33]。综合这些研究结果可以推测,驴乳具有较强的抗氧化功能可能归因于Nrf2-ARE通路的激活。
2.2.3 驴乳可能通过过氧化物酶体增殖物激活受体家族(PPARs)信号通路调节机体的抗氧化功能
PPARs主要包括PPARα、PPARβ和PPARγ 3种核转录因子,在脂肪代谢和炎症反应中发挥重要作用。其中,PPARα的激活可增加酶活性和线粒体氧化,从而激活机体的抗炎和抗氧化功能[34]。PPARα的激活剂吡格列酮可上调SOD1、GSH-Px1、CAT和解偶联蛋白2(UCP2)的基因表达,进而提高小鼠耳蜗细胞的抗氧化能力[35]。在驴乳的研究中指出,驴乳三酰基甘油的sn-2位置上具有较高的棕榈酸含量[36-37];而sn-2位的棕榈酸可以调节机体组织中N-乙酰乙醇胺含量,N-乙酰乙醇胺又是PPARα的激活剂[38]。因此,驴乳具有较强的抗氧化功能可能与其对PPARs信号通路的调节作用有关,但确切的机制还有待于进一步探讨。
3 驴乳的其他生物学功能
驴乳除了具有抗氧化活性外,还具有抗炎、抗病毒和抗菌等生物学功能。Yvon等[39]研究发现,驴乳可以促进患回肠炎小鼠内源性抗菌肽的分泌而发挥抗炎作用,从而减少小鼠肠道菌群失衡。Trinchese等[40]和Lionetti等[6]饲喂小鼠驴乳,结果表明,驴乳可以降低小鼠血清肿瘤坏死因(TNF-α)、白细胞介素-1(IL-1)、白细胞介素-10(IL-10)和脂多糖(LPS)含量。另外,Tafaro等[37]研究发现,驴初乳可介导人淋巴细胞的免疫球蛋白A(IgA)应答,而常乳则更多地介导免疫球蛋白G(IgG)应答。
4 小 结
综上所述,驴乳具有清除自由基和提高动物机体抗氧化能力的效果。本文根据前人研究综合总结了驴乳可能通过PPARs和Nrf2-ARE信号通路进行抗氧化调节,发挥驴乳的抗氧化功能。但目前相关的研究尚且鲜见,有必要深入探讨驴乳和驴乳各成分的抗氧化功能及其作用机理,为驴乳的开发提供理论基础。
