CDR1as在前列腺癌中表达及作用机制探索
2021-08-03张春雷杨琦周逢海徐东波康亚芬常德辉
张春雷 杨琦 周逢海 徐东波 康亚芬 常德辉
CDR1as(又称ciRS-7,hsa_circ_0001946)已被证明在包括肝细胞癌、肺癌、胰腺癌、卵巢癌等[1]在内的多种肿瘤疾病中发挥促进肿瘤进展的作用,在膀胱癌中通过CDR1as-miR-7-p21/CDR1as-miR1270途径发挥抗肿瘤作用[2]。CDR1as是目前研究广泛及功能强大的环状RNA(circular RNA, circRNA),然而其在前列腺癌发生、发展中的作用目前尚无论证,我们通过对34例前列腺癌患者术后标本的癌及癌旁配对组织样本进行高通量测序,发现CDR1as在前列腺癌组织中显著低表达,说明CDR1as可能在前列腺癌的疾病发生及进展中发挥重要作用,对此,我们通过基因检测结合数据库分析,探索CDR1as在前列腺癌中可能的作用及机制。
材料与方法
一、前列腺癌患者组织标本获取及测序
选取就诊于中国人民解放军联勤保障部队第九四〇医院术后病理诊断为前列腺癌患者的肿瘤组织及癌旁组织(已通过医院伦理审核,审批编号:2020KYLL030),一部分留作提取RNA,一部分送至测序公司进行测序,差异分析采用edgeR分析方法,差异表达标准为Fold change>2.0 且P值<0.05,细胞测序标准与之相同。
二、细胞培养
将细胞接种于含有4 ml(10%胎牛血清)培养液的培养瓶中,放于37 ℃、5% CO2条件下的培养箱中进行培养。RWPE1细胞培养在PEpiCM培养液中,LNCap细胞培养在RPMI-1640培养液中,PC3细胞培养在F-12(ham)培养液中,培养液每2天更换一次,细胞在生长到70%~80% 时用0.5 ml 0.25% 的胰蛋白酶/ EDTA室温下消化并传代。
三、siRNA干扰处理
常规消化细胞并计数,在6孔板中预铺1×105个细胞,1 d后至生长对数期,使用预热的OPTI-MEM配置转染试剂悬液和10 mmol/L干扰RNA(siRNA)悬液,按照说明书将混匀的转染试剂溶液滴加到培养液中,放入培养箱中继续培养。siRNA1:GCACCTGTGTCAAGGTCTTTT;siRNA2:GGT-CTTCCAGCGACTTCAATT;siRNA3:GTGCTGATCTTCTGACATTTT。
四、RNA提取
将组织在液氮中磨碎,用匀浆仪进行匀浆处理后采用Trizol法常规提取RNA;将培养贴壁细胞用PBS清洗2遍,之后采用Trizol法常规提取RNA。
五、反转录及RT-qPCR
按照反转录试剂盒(PrimeScript®RT Master Mix试剂,Takara,中国)提供的标准方法进行配置,放至PCR仪,按照37 ℃ 15 min、85 ℃ 5 s条件进行反转录;按照RT-qPCR试剂盒(SYBR®Premix Ex TaqTM试剂,Takara,中国)提供的标准方法进行配置,放入仪器中,采用两步法RT-qPCR的默认设定程序进行检测,每个样品做3次重复。引物如下:CDR1as(F:CGTCTCCAGTGTGCTGATCT;R:GTCCGGAAGACATGGATT),β-actin(F:CGC-GAGAAGATGCCCAGATC;R:TCACCGGAGT-CCATCACG)。
六、CDR1as作用机制的预测
采用miRanda软件预测CDR1as与miRNA结合位点,利用癌症基因组图谱(The Cancer Genome Atlas, TCGA)数据库中的临床数据信息,分析差异表达的基因对前列腺癌预后的影响,通过circinteractome网站(https://circinteractome.nia.nih.gov/)预测CDR1as是否与蛋白质存在结合位点,利用ORF Finder网站(https://www.ncbi.nlm.nih.gov/orffinder/)预测CDR1as是否拥有开放阅读框(open reading frames, ORF)及IRESite网站(http://iresite.org/IRESite_web.php)预测是否具有内在的核糖体进入结合位点(internal ribosome entry site, IRES)。
七、统计学方法
采用SPSS 21.0软件分析数据,计量资料采用均数±标准差表示,两组间比较采用t检验,采用Pearson相关性分析预测基因与CDR1as表达的相关性及利用生存分析进行预后分析,癌与癌旁组织中表达差异变化采用表达量差值进行分析(T-N),P<0.05为差异具有统计学意义。
结 果
一、CDR1as在前列腺癌患者组织及前列腺癌相关细胞系中的表达情况
在34例前列腺癌患者术后标本的配对样本中,CDR1as在癌组织中表达量显著低于癌旁组织(P<0.05,图1A);为验证测序结果的准确性,选取17例前列腺癌患者术后标本的癌及癌旁配对组织样本进行RT-qPCR验证,结果表明,在3组配对的组织样本中,CDR1as在癌组织中的表达量高于癌旁组织,在其余的14组配对组织样本中,癌组织中的表达量均低于癌旁组织,CDR1as在癌组织中的表达量低于癌旁组织,差异有统计学意义(P<0.05,图1B、1C)。相比于前列腺正常上皮细胞系RWPE1,CDR1as在前列腺癌细胞系PC3中高表达,在LNCap中低表达(P<0.05)。
二、筛选被CDR1as调控的下游靶基因
为筛选在前列腺癌发生、发展中可能被CDR1as调控的下游靶基因,采用3种CDR1as的siRNA对表达量升高的PC3细胞进行处理,模拟肿瘤组织中CDR1as表达下调,收集处理过的细胞进行mRNA测序,取3组处理细胞测序结果中同时上调或下调的mRNA,并选取与组织测序结果中差异表达mRNA变化趋势一致的mRNA。结果表明经过siRNA处理后,3组PC3细胞中的CDR1as表达量均下调(图2A),与对照组相比,测序的3组配对样本中共有225个显著上调的mRNA和228个显著下调的mRNA,组织测序结果中共有401个显著上调的mRNA、1 010个显著下调的mRNA,共筛选出7个显著上调的mRNA和37个显著下调的mRNA作为在前列腺癌发生及发展中被CDR1as调控的下游靶基因(图2B)。

A:34例前列腺癌患者术后标本的癌及癌旁配对组织样本中CDR1as测序表达量对比;B、C:17例前列腺癌患者肿瘤组织与癌旁组织CDR1as表达量的RT-qPCR验证;D:CDR1as在3种前列腺癌相关细胞系中的表达图1 CDR1as在前列腺癌组织及细胞中的表达量(*P<0.05,**P<0.01)
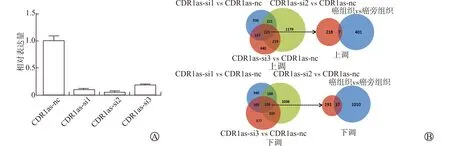
A:PC3细胞中采用3种siRNA干扰处理后CDR1as表达量;B:筛选组织测序结果及PC3细胞采用3种siRNA干扰处理后测序结果中差异表达变化趋势一致的mRNA 图2 被CDR1as调控的下游靶基因的筛选
三、探索CDR1as与差异表达基因的相关性
采用Pearson相关性分析组织测序结果, 探索肿瘤组织中与CDR1as表达变化存在相关性的基因,并对这些基因在癌与癌旁组织中表达差异变化与CDR1as在癌与癌旁组织中表达差异变化(用表达量差值表示)是否存在相关性进行检测,表1列举了与前列腺癌或其他恶性肿瘤发生及发展存在相关性的基因,共有19个基因在前列腺癌患者肿瘤组织中与CDR1as表达变化存在相关性,其中表达上调的为BIRC5和PRDX4,表达下调的为RASGRP2、MRVI1、RUNX1T1、CD200、RARRES2、ASPA、CD69、RGS22、GSTM5、LRP4、ASXL3、MYO1F、CCL5、FBLN2、PRRX2、ACOX2和INPP5D;共有12个基因在癌与癌旁组织中表达差异变化与CDR1as表达差异变化存在相关性,其中表达上调的为PRDX4,表达下调的为MRVI1、RUNX1T1、CD200、RARRES2、ASPA、GSTM5、LRP4、FBLN2、PRRX2、ACOX2和INPP5D(P<0.05)。采用TCGA数据库中的临床数据信息,分析差异表达的基因对前列腺癌预后的影响,结果表明低表达的GSTM5与前列腺癌不良预后有关(P<0.05,图3)。

图3 GSTM5表达对前列腺癌患者预后分析

表1 在前列腺癌患者组织中与CDR1as表达变化趋势存在相关性的mRNA
四、预测CDR1as与miRNA结合位点
为进一步探索CDR1as在前列腺癌发生及发展中可能的作用机制,采用miRanda软件预测CDR1as与miRNA结合位点,共检测出CDR1as与68个miRNA存在结合位点,存在5个以上结合位点的miRNA为hsa-miR-7-5p、hsa-miR-3617-5p、hsa-miR-8056、hsa-miR-641、hsa-miR-1290、hsa-miR-1246、hsa-miR-3156-5p、hsa-miR-139-3p及hsa-miR-3167(图4)。

图4 miRanda软件预测CDR1as与miRNA结合位点
五、预测CDR1as是否与蛋白质相互作用或翻译蛋白的可能性
circRNA可以与蛋白质相互结合或翻译蛋白而发挥作用,通过circinteractome网站预测CDR1as是否与蛋白质存在结合位点,结果表明CDR1as序列与蛋白质无结合位点,反向剪接位点与IGF2BP2存在结合位点,其侧翼序列与FUS、IGF2BP2、IGF2BP3存在结合位点(表2)。为探索circRNA是否具有翻译蛋白质的潜能,利用ORF Finder网站及IRES网站进行预测,结果显示CDR1as具有14个ORF(表3)及8个IRES(表4)。

表2 CDR1as与蛋白质结合位点预测

表3 CDR1as ORF预测

表4 CDR1as IRES预测
讨 论
CDR1as具有1 485个碱基,它是由CDR1反义链环化形成的circRNA,其序列具有丰富的miRNA结合位点,不但种类多,而且与多个miRNA具有5个以上的miRNA结合位点,预示着其miRNA海绵作用机制功能强大,miRanda软件预测其与miRNA结合位点证明了这一点。通过对34例前列腺癌患者术后标本的癌及癌旁配对组织样本测序结果发现CDR1as在前列腺癌组织中显著低表达,CDR1as在另外17例前列腺癌患者的癌及癌旁配对组织标本中的RT-qPCR表达量也验证了测序结果的准确性,表明其可能在前列腺癌的发生、发展中发挥作用。为了进一步探索其潜在作用和可能的作用机制,我们通过测序及数据统计分析方式筛选出可能被CDR1as影响表达的候选基因,根据候选基因的功能推测CDR1as在前列腺癌疾病中的作用。PRDX4是筛选出的与CDR1as在前列腺癌患者组织中变化相关且在前列腺癌组织中上调的基因,前列腺癌组织中PRDX4过表达,并促进前列腺癌细胞的增殖[3],但下调的CDR1as如果通过miRNA海绵机制发挥作用,理论上PRDX4在前列腺癌组织中受到miRNA作用会出现下调,说明CDR1as可能通过其他途径调节PRDX4,比如与长链非编码RNA等相互协调共同发挥作用。其他与CDR1as变化显著相关的几个基因主要发挥抑癌作用,比如miR-940通过下调MRVI1促进子宫内膜癌的增殖和转移[4],RUNX1T1抑制结直肠癌细胞的增殖并促进化疗药物抵抗[5],ASPA、PRRX2和GSTM5在前列腺癌组织中显著下调[6-7],FBLN2在鼻咽癌中发挥抗血管生成和肿瘤抑制作用[8],ACOX2在心脏恶性肿瘤中表达缺乏[9],INPP5D作为抑癌基因p53的下游靶基因,在多个肿瘤中的缺氧诱导调控过程起到促进细胞凋亡的作用[10],而且低表达的GSTM5与前列腺癌不良预后有关。如果前列腺癌肿瘤组织中低表达的CDR1as导致这些发挥抑癌作用的基因表达降低,那么其发挥促肿瘤作用的机制可能是通过这些基因实现的,这些基因表达的下调也证明CDR1as的表达下降可能是前列腺癌疾病进展的因素。
肿瘤微环境是调节肿瘤疾病进展的重要因素,而免疫调节在肿瘤微环境中发挥重要作用。在与CDR1as表达变化显著相关的基因中,RARRES2表达的蛋白是一种内源性白细胞趋化剂,可通过其受体ChemR23募集先天免疫细胞。在肿瘤微环境中增强RARRES2可以部分抑制免疫效应细胞的募集,从而显著抑制肿瘤的生长,升高的RARRES2可以通过增加PTEN的表达及降低PD-L1的表达显著抑制前列腺癌细胞系DU145的成瘤作用[11],说明 RARRES2表达的下调也是前列腺癌进展的可能因素之一。另外,肿瘤特异性TH1细胞在抗癌免疫反应中也起到重要作用,CD200和CD69是肿瘤特异性CD4+T细胞表面特异性分子[12],两者表达的下降是否意味着下调的CDR1as也影响着肿瘤特异性T细胞的表达,进而影响其抗肿瘤的功能,值得进一步探索。
当然,这些与CDR1as表达密切相关的下调基因也有发挥促癌作用的基因,比如LRP4促进胃癌及甲状腺癌的发展[13-14],FBLN2在肺腺癌中也是恶性进展的促进因子[15]。那么它们的下调意味着作为抑癌因素存在。此外,ASXL3、MYO1F和CCL5也是促癌基因[16-18],尤其是有报道表明CCL5通过激活β-catenin/STAT3信号通路促进前列腺癌干细胞的转移[19],但这些基因在肿瘤组织与癌旁组织中差异变化并未体现出与CDR1as表达差异变化存在相关性,说明它们可能并非CDR1as调控。总体来说,这些与CDR1as表达密切相关的基因主要以发挥抑癌作用的基因为主。
除了miRNA海绵作用,circRNA还可以通过与蛋白质相互作用,甚至翻译蛋白发挥作用。虽然CDR1as是反义链circRNA,但如果其具备翻译蛋白质的必备条件,也有可能通过翻译成蛋白质发挥作用。已有研究表明CDR1as通过破坏P53/MDM2复合物从而限制P53蛋白的泛素化来促进其稳定性,最终达到抑制胶质瘤的进展[20],因此CDR1as在前列腺癌中也可能通过与蛋白质相互作用的机制发挥功能。通过数据库的预测及分析,CDR1as具有14个ORF及8个IRES,那么其能否翻译蛋白质也是值得探索的。另外,CDR1as的剪接位点及侧翼序列可能与FUS、IGF2BP2、IGF2BP3相结合,而FUS/TLS是前列腺癌细胞中雄激素受体的共激活因子,可以促进前列腺癌细胞系LNCap细胞的雄激素依赖性增殖[21],IGF2BP3在前列腺癌组织中高表达,且与高Gleason评分相关[22],下调的CDR1as可能因为对FUS、IGF2BP2、IGF2BP3抑制作用减轻而发挥促肿瘤作用。
在进行前列腺癌细胞系的CDR1as表达量验证时发现,前列腺癌骨转移细胞系PC3中CDR1as高表达,在LNCap细胞中低表达,考虑原因可能是细胞系为纯化的细胞,导致其分子表达量与组织中表达量不一致,另外PC3细胞系并不能很好的作为前列腺癌研究模型,PC3细胞为骨转移细胞系,其基因表达与原位细胞系的基因表达差异比较大,但是CDR1as在骨转移细胞系中的高表达是否意味着随着表达量的改变导致转移的发生也是将要探索的方向。这种现象也提示我们在研究CDR1as在前列腺癌疾病进程中的作用应选择LNCap细胞模型。总体而言,通过对CDR1as在肿瘤组织中低表达以及CDR1as与差异表达基因的相关性进行统计分析,得出低表达的CDR1as可能促进前列腺癌的发生、发展的结论,其具体功能有待进一步进行细胞生物学的鉴定,比如利用CDR1as低表达的LNCap细胞构建CDR1as过表达的稳转株并检测其生物学变化,以及CDR1as精确的作用机制等都有待进一步探索。
