继发性肺结核致病机制的再认识及索状因子在其中的作用
2021-07-09刘镇铭艾沁悦耿雪薇王嘉军石廷玉
刘镇铭,艾沁悦,耿雪薇,黄 胜,王嘉军,石廷玉
湖北民族大学医学部基础医学院病原生物学教研室,湖北恩施 445000
结核分枝杆菌(Mycobacteriumtuberculosis,MTB)感染目前仍是紧急的全球健康问题,2018年约有1000万人患结核病和150多万人死于结核病,中国新增病例数居印度之后为全球第二。耐多药结核仍是公共卫生危机和健康安全威胁,据世界卫生组织统计,全球大约1/4的人潜伏感染MTB,具有5%~15%的风险患病。对于免疫受损的患者,如人类免疫缺陷病毒(human immunodeficiency virus,HIV)感染者、营养不良、糖尿病患者或吸烟患者风险更高。MTB和HIV的双重感染以及耐多药结核菌的出现加剧了问题的严重性,并阻碍了防控的效果。缺乏对MTB继发感染致病机制的认识是阻碍疫苗和药物研发的主要原因。本文总结了原发性感染和继发性感染的过程,并对目前广泛接受的继发性感染致病机制存在的问题及产生的原因,以及致病机制新的认识及索状因子的作用进行了阐述,以促进对MTB继发感染致病机制的理解,为疫苗研发和药物开发提供新的认识。
原发性感染和继发性感染
MTB的感染周期经历了感染个体并诱导其产生治愈损伤的适应性免疫(原发感染);持留几十年之后,通过活化或再感染发展成为活动性肺结核,在肺部产生空洞,并在空洞中大量繁殖,最后通过咳嗽进入环境感染新的个体(继发感染)。大量MTB通过空气进行传播,该过程可以持续数十年。有研究显示,在对1例晚期肺结核患者痰液中的MTB进行连续监测后发现,24 h内产生的细菌数量达15~40亿[1]。由于宿主的疾病或死亡可减少MTB传播的机会,因此大量无症状携带者是MTB重要的传染源。原发性感染可通过产生有效的适应性免疫,保护宿主不会因为感染而死亡。继发性感染让MTB逃避和破坏宿主的系统免疫产生空洞,以进行增殖和传播。
原发性感染是个体首次感染MTB[2]。虽然原发性感染可能会发展为脑膜炎或播散性结核,尤其是在年幼或免疫抑制的个体,但原发性感染通常发展和扩散到局部淋巴结形成干酪样肉芽肿,几周后由于适应性免疫的产生而逐渐消退。虽然损伤可能治愈,但MTB仍然存在。由于有大量的原发性感染动物模型和人类组织进行研究,因此在理解原发性感染的特征性损伤—干酪样肉芽肿的发展过程和作用方面已取得很大进展[3-4]。
继发性感染发生于已经产生适应性免疫的个体[5],在很多方面不同于原发性感染,如:对宿主敏感性的调节、临床表现、并发症和年龄分布等方面[6]。继发性感染通常发生于肺上叶,并不涉及淋巴结或其他器官。大约90%的个体不需要治疗而能够自愈。不能自愈的患者并不是免疫力不足,因为除部分肺之外,患者具有足够的免疫力防止全身性感染。事实上,死于急性继发性感染的个体通常是15~40岁具有免疫活性的个体[7]。继发性感染不仅不能让宿主产生适应性免疫,反而是自愈或通过治疗康复的个体具有更高的风险再次发病[8]。
目前继发性感染致病机制存在的问题
目前认为,干酪样肉芽肿是原发性感染和继发性感染的共同特征,而空洞的产生是由干酪样肉芽肿侵蚀到支气管所形成的[9]。一个科学理论的价值在于其是否能够很好地解释自然现象,继发性感染的致病机制应该能够解释现存问题,然而目前普遍接受的继发性感染的致病机制不能回答下述问题并与一些事实不符。
目前致病机制不能回答的问题MTB是如何在宿主其他部位存在有效免疫的情况下,存在于肺的特定部位?各种肺损伤如何独立发生于同一个体的肺中,而在其他部位好像并不存在?95%的个体能够自愈的原因是什么?为什么继发性感染康复的个体具有更高的风险再次发病[8]?超敏反应在继发性感染中扮演了什么角色?不像其他病原体如HIV通过修饰抗原来逃避免疫反应,而MTB反而保持其抗原非常的稳定,甚至比一些重要的结构基因都更少改变[10]。为什么MTB需要这些抗原表位?不能回答这些问题就不能使人相信能够通过疫苗阻止结核的传播。缺乏对MTB潜伏感染的免疫和致病机制的理解被普遍认为是阻碍疫苗和药物研发的主要原因。如果对分枝杆菌感染产生保护性免疫的特征不理解,没人能够合理的设计或测试疫苗。
与目前致病机制不符的事实以下的一些描述来源于对成百上千例患者的观察结果,应该属于事实:(1)发展中的继发性感染首先是特征性支气管阻塞,然后产生继发阻塞性肺炎,最终产生空洞[11]。(2)空洞是由干酪样肺炎破碎后形成的,而不是肉芽肿侵蚀产生的[12]。(3)肉芽肿是对干酪样坏死的反应而不是导致干酪样坏死的原因[12]。(4)伴有空洞产生的干酪样肺炎通常含有非常少的MTB[13]。(5)MTB抗原在具有非常少的抗酸杆菌区域的细胞内聚集[14]。
目前致病机制产生的原因
结核病的研究起始于1800年左右,到20世纪50年代,由于抗生素的使用人们对MTB研究的兴趣开始下降[1]。直到20世纪80年代结核病的再次出现,才恢复了新一代研究人员对结核病的兴趣。因此,对结核病的研究出现了数十年的中断,前后几乎没有任何信息传递。处于前抗生素时代时,研究对象是未经治疗的患者。几乎每一名研究人员都研究了上千例的继发性感染患者,他们认为MTB是通过支气管扩散并产生肺炎,最终产生了干酪化和空洞[13]。当时很多研究人员已经认识到,结核病具有增生性和渗出性两个不同的病理过程[8,13]。增生性过程是指现在所说的结节性结核或肉芽肿,渗出性过程指的是肺炎。增生性阶段是结核病的原发性感染阶段,渗出性阶段是结核病的继发性感染阶段。因此,当时的观点认为继发性感染是由肺炎而非由肉芽肿引起的[1]。
空洞产生于肉芽肿向支气管侵蚀的观点形成并不久。当耐药性MTB和HIV的出现重新激起人们研究结核病的兴趣时,需要寻求一个人类结核病的研究模式,但新一代研究人员很难获得未经治疗的人类结核病肺组织,因此缺乏对未经治疗的人类肺组织的认识。由于牛分枝杆菌能够导致兔的肺组织产生空洞,其空洞是肉芽肿对支气管侵蚀所产生的[15]。在缺乏其他观点的情况下,肉芽肿是人类原发性感染和继发性感染的主要损伤模式观点就形成了。这对于牛分枝杆菌产生空洞而言是准确的,而对于MTB产生空洞的情况是不恰当的[16]。因为在MTB感染人之后形成的肉芽肿都非常小,不足以对支气管侵蚀产生空洞[12]。再则,牛分枝杆菌不会产生继发性感染,而是通过具有很强攻击性的原发性感染产生干酪样肉芽肿,侵蚀咽、支气管、胸膜、肠道、泌尿道和乳腺等组织传播病原菌[17]。这不同于人类MTB通过继发性感染产生空洞的方式传播病原菌。由于没有认识到牛分枝杆菌和MTB造成损伤方式的差异,因此干酪样肉芽肿是原发性感染和继发性感染共同特征性损伤的概念成为基本模式,伴随免疫学、分子微生物学和遗传学的兴起,指导了结核病的研究。
对结核病致病机制新的认识
早期继发性感染继发性感染开始时,毒性MTB通常存在肺尖部位,虽然该个体具有足够的免疫力能够自愈干酪样肉芽肿并阻止身体其他任何部位的感染。对原发性感染和继发性感染分布情况的比较显示,原发性感染随机分布在肺部,而继发性感染主要分布在肺尖[13]。因此有研究者提出,为什么继发性感染对肺尖部分更敏感[8]。通常认为MTB优先在肺尖部位生长是因为肺尖的氧分压高[18],然而该观点未获得认同。1983年,den Boon等[8]就质疑了高的氧分压导致结核定位于肺尖的假设,他们认为淋巴瘀滞和抗原物质不能清除是MTB在肺尖定植的原因。肺尖部分是肺最独立的一个区域,因为其具有最低的通气量、血液灌注和淋巴流。很多证据支持这种假设,低血流是早期继发性感染致病机制的一个因素。首先,卧床休息在抗生素治疗问世前是最好的治疗措施之一[19]。卧床休息增加了肺尖的血流,尤其是将脚抬高效果更好。其次,位于空气稀薄的高海拔地区患者因为需要更剧烈的呼吸作用,通常会改善肺结核。最后,因肺二尖瓣狭窄而具有高肺动脉压的结核病患者很少,而由于肺动脉瓣狭窄导致肺动脉压降低的患者会遭受MTB更大的损伤[18]。
由于测量肺各部分呼吸效果技术的成熟,如四维CT扫描能够在三维空间看到肺在不同时间的形态[20]。新技术的应用发现肺的各个部分运动是不一样的,在隔膜运动的底部运动最大,胸廓运动的胸下部运动次之,在肋骨固定的肺尖部位最少。当直立和侧卧时,在肺尖部位的通气也是非常少的,因为肺并没有舒张和参与呼吸作用。这表明继发性感染起始于肺尖部位是因其不运动、低通气量和低血流灌注,从而导致了淋巴瘀滞和抗原物质不能清除,最终导致了支气管阻塞。
支气管阻塞产生内源性脂质性肺炎理解继发性感染致病机制的一个重要线索为支气管阻塞是继发性感染早期阶段的一个特征。细支气管阻塞导致继发感染[21]。Medlar等[12]提出将继发性感染称为支气管原结核,因为其通过支气管扩散从而产生阻塞和结核性肺炎。目前研究也发现,早期继发性感染产生特征性X射线表现为树芽征,并通常伴随附近肺的磨玻璃影,提示有内源性脂质性肺炎。放射影像显示肺结核通常呈现出上叶楔形病变,楔形的顶点就是阻塞的支气管[13]。组织病理学分析表明,小叶中心的“树”型损伤是由于支气管末端、细支气管或肺泡管阻塞所导致。“芽”是由阻塞导致的炎症病灶点。小叶中心“树”型损伤可能再生长合并成为既有干酪样坏死也有周围局灶性炎症的小叶实变。空洞产生于小叶中心,几个小叶中心形成的空洞逐渐合并成为更大的空洞。
异物或肿瘤导致的支气管阻塞也能导致阻塞性肺炎,类似于继发性感染导致的损伤[22]。阻塞性肺炎是内源性脂质性肺炎,由于大量脂质聚集而成黄色又称为金黄色肺炎[23]。大量的脂质来源于2型肺细胞产生的表面活性剂,通常能够通过支气管清除。如果支气管阻塞,表面活性剂就会被肺泡巨噬细胞吞噬并进行化学修饰,随着脂质成分增加逐渐形成泡沫巨噬细胞[24]。MTB似乎可选择性产生支气管内感染,从而导致阻塞,支气管阻塞又造成聚集脂质的泡沫巨噬细胞聚集在肺泡中。Betancourt等[23]提出支气管阻塞是导致宿主局部免疫应答受到抑制的重要原因,对继发性感染的发展起到非常重要的作用。缓解阻塞会导致结核消退[25]。总之,这些结果支持支气管阻塞是继发性感染致病机制的重要原因之一。由于继发性感染具有阻塞性肺炎的多种典型特征,因此对阻塞性肺炎的研究将为继发性感染致病机制的研究提供线索。
继发性感染起始于内源性脂质性肺炎现有大多数文献将继发性感染描述为宿主与MTB的战争,即巨噬细胞被募集激活以控制和杀死MTB,MTB试图增殖并杀死巨噬细胞,双方力量强弱对比决定了感染的结局。这对于原发性感染来说是合理的,而对于继发性感染并非如此。多项研究显示,继发性感染是以脂质性肺炎开始的[1,13,15,26]。
继发性感染是由潜伏的MTB复活或新感染的MTB所导致,发生于已产生适应性免疫的个体,损伤起始于内源性脂质性肺炎[27],而不是由于大量MTB增殖造成的或肉芽肿侵蚀到支气管产生空洞。少部分MTB将部分肺组织隔离出来,同时安静地聚集MTB抗原和宿主脂质成分,这些活性成分聚集起来达到一定程度时引起的内源性脂质性肺炎触发了突然的干酪样坏死,从而发展成为空洞或纤维干酪样结核[28]。在干酪样肉芽肿形成之前的损伤是完全可逆和无症状的,但是一旦形成干酪样肉芽肿之后损伤是不可逆和高度毒性的。
索状因子在致病机制中的作用
Hunter等[29]研究认为,索状因子海藻糖-6,6’-二分枝菌酸(trehalose-6,6’-dimycolate,TDM)是理解MTB导致内源性脂质性肺炎触发干酪样坏死产生这个现象的关键。TDM是毒性MTB产生最多的脂质成分,也是长期以来是最为神秘的物质,因为当TDM与油性溶剂混合后,会从无毒性变成高毒性。
索状因子的生物学特性Middlebrook等[30]于1947年报道,毒性MTB具有独特的表型,即蛇纹索状结构。Bloch[31]认为蛇纹索状结构的形成与MTB细胞表面的脂质成分有关,他采用温和的有机溶剂提取了细菌表面的脂质成分,结果发现提取后的细菌仍具有活性,但不能形成索状表型,因此将这种脂质成分被称为索状因子。这种脂质成分就是TDM[32]。TDM由1个海藻糖和2个长链脂肪酸形成,存在于细胞壁,形成很厚的荚膜导致MTB以索状生长,是MTB重要的毒性物质。无毒的MTB也具有TDM,但其表面游离的非常少[33]。TDM完全不溶于水,当细菌释放出细胞后就留在其表面,因此可以通过温和的有机溶剂如石油醚,在不杀死细菌的情况下将细胞表面的TDM提取出来,通过这种方法可以得到几乎纯净的TDM[34]。TDM是唯一一种在不杀死细胞的情况下可以大量获得的脂质成分[32]。
Retzinger等[35]研究发现,TDM在空气-水、塑料-水和油-水界面的行为类似,能够自发形成结晶单层分子,由此推测TDM可能是在空洞中的空气界面形成单层分子导致毒性激活,维持空洞产生。TDM与脂质相互作用是MTB致病机制的核心基础[29,36]。TDM具有多种不同的生物学活性,其分子很大,覆盖在细菌表面,不溶于水,因此可阻止吞噬溶酶体融合和吞噬体酸化,从而阻止微生物被巨噬细胞杀死和抗原提呈[37]。TDM的独特性在于其生物学活性与溶剂相关。在生理盐水中,TDM是无毒的,对老鼠的半数致死剂量(50% lethal dose,LD50)超过50 000 μg。然而,当TDM以单分子层存在于脂质微滴表面时,具有完全不同的物理结构和生物学活性[38](图1A),其能够在油-水界面自发形成二维的单分子层,这是已知的最坚硬的脂质单分子层[39]。这种单分子层能够在任何疏水界面自发形成。当毒性MTB与脂质微滴接触时,它们立即进入脂质微滴,释放出的TDM在水-脂质界面形成单分子层构象(图1A)。TDM的单分子层构象很稳定,在老鼠中以单分子层存在的半衰期为4 d[40]。以单分子构象存在时,TDM的毒性很强,LD50为30 μg,能在小鼠体内产生异物肉芽肿,在免疫小鼠体内产生过敏性肉芽肿[41]。每天注射10 μg的TDM乳浊液可导致C57BL1/6小鼠发生出血性肺炎、恶病质和死亡。更大剂量(50~100 μg)的TDM乳浊液会立即导致死亡。TDM与油的混合物是增强和刺激免疫反应最有效的佐剂[42]。MTB细胞表面的TDM可保护细菌不被巨噬细胞杀死。然而,从1个细菌表面提取的TDM要是以单层分子形式存在,能够在数分钟之内杀死20个巨噬细胞[29]。这不仅对巨噬细胞如此,其他与TDM单层分子接触的细胞也会死亡。
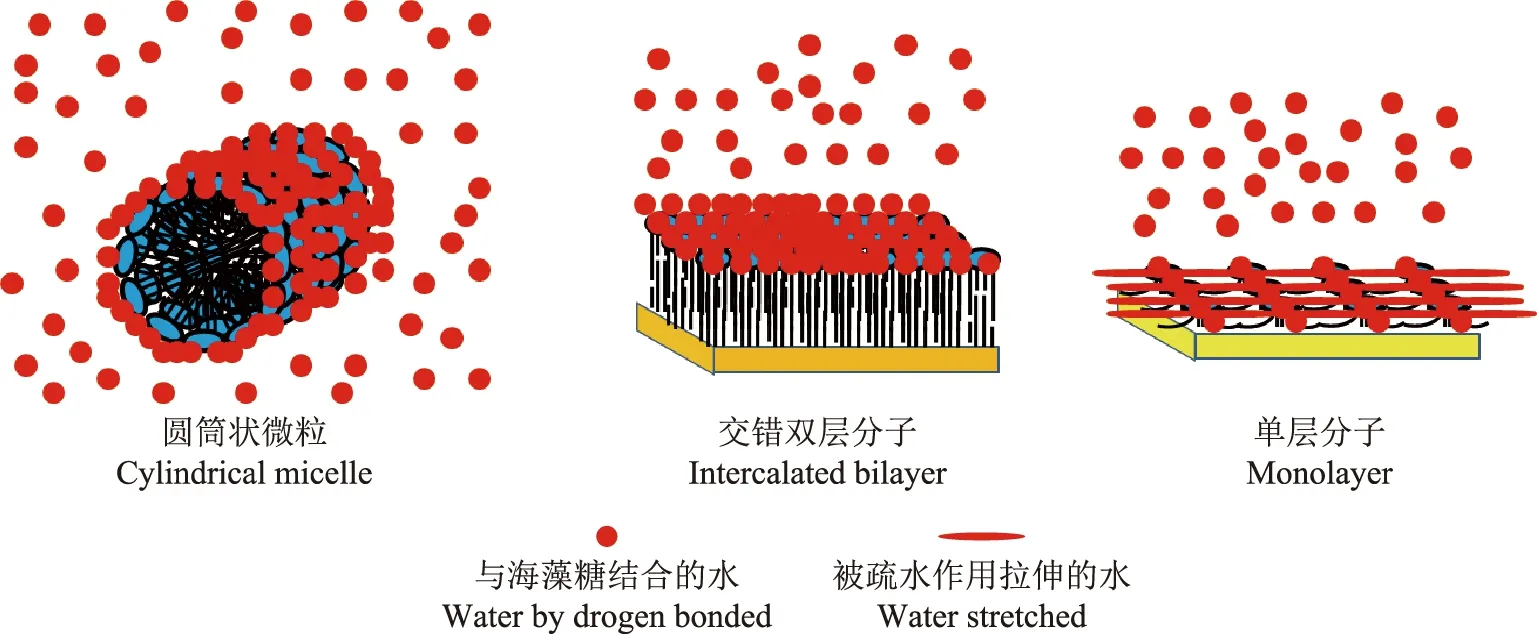
Hunter等[33]研究认为,根据其存在方式,TDM具有完全不同的生物学活性,这些生物学活性与TDM和水分子之间的生物物理作用有关。在水环境中,TDM形成圆筒状的微粒、在分枝杆菌细胞壁形成的交错双层分子和在水-疏水界面形成的分子单层(图2)[39]。微粒和双层的表面完全是由海藻糖构成,结构非常稳定,因为长链分枝杆菌酸能有效地包裹在一起。
不同于圆筒状的微粒或在分枝杆菌细胞壁形成的交错双层分子,TDM单层表面不完全是海藻糖分子所形成。海藻糖仅占30%,其余70%的表面是由暴露的分枝菌酸链以严格的阵列排列。这种构象比其他任何已知生物来源的双亲分子形成的单层分子都要坚硬和稳定[35],与毒性矿物晶体如石英和鳞石英有物理和生物学类似性,表明其毒性具有共同的作用机制[36]。
虽然学术界对TDM的这些生物学活性已有充分认识,但由于TDM的毒性需要脂质的存在,从而认为这样的致病条件是“非生理的”,因此,一直以来学术界并不认同TDM是肺结核毒性因子[43]。然而最新研究结果表明,与脂质相互作用激活TDM毒性是内源性脂质性肺炎触发干酪样坏死的重要原因[29]。
索状因子是干酪样肺炎的触发因子导致TDM触发干酪样肺炎发生的可能原因是构象改变,从而刺激大量肿瘤坏死因子快速释放。Geisel等[44]研究证实,TDM颗粒越大诱导巨噬细胞释放肿瘤坏死因子能力越强。在直径为90 μm小球上,TDM诱导巨噬细胞释放肿瘤坏死因子达2600 pg/ml;而在1 μm直径小球上,TDM只能诱导100 pg/ml的肿瘤坏死因子产生。Goren[45]也证实,注射TDM与油的混合物比注射相同量TDM与水的混合物毒性更强。Hunter 等[13]研究发现,包被TDM的小球能够被巨噬细胞吞噬,形成肉芽肿。相反,包被在平面上或大球表面的TDM可导致巨噬细胞黏附、扩散和快速裂解[36]。
继发感染分离出一部分肺以促进宿主脂质成分和MTB抗原的聚集,为干酪样肺炎和空洞的产生创造条件。在阻塞性肺炎中,脂质成分的逐渐聚集导致脂质微滴大小逐渐增加。干酪样肺炎的触发或许是包被有TDM的脂质微滴大小增加到临界值的反应,并激活大量肿瘤坏死因子释放,从而触发了突然的干酪样肺炎,然后快速产生干酪样坏死,导致空洞产生。
空洞产生一旦肺损伤发展成为干酪样肺炎,有可能软化并破碎,以至于部分肺被咳出形成空洞[1],也有观点认为空洞的产生是由于坏死的肺基质被蛋白酶降解后所产生[46]。刚产生的空洞是粗糙的,随着时间推移,在空洞表面逐渐形成了一层薄薄的纤维,几乎没有炎症,细菌只在腔的内表面生长。有研究发现这种空腔总是集中在支气管上,其内表面是一种光滑的蜡状物质,最后进入支气管[13]。Hunter等[29]研究发现,其内表面是以菌膜生长的大量微生物。在体外,MTB在没有表面活性剂的液体培养基中培养时就优先以菌膜形式生长,并促使其穿过表面并攀爬到容器侧面[13]。这种特性与索状因子单层分子的刚性有关[29]。
空洞维持空洞一旦形成,通常很难愈合,并不断产生毒性MTB排到环境中。在空洞中,大多数MTB存在于细胞外液体中。很少有人去解释这种现象。在空洞中产生的MTB数量非常巨大。中度肺结核患者每天产生的细菌数量为14~40亿[29]。空洞很难治愈,除非这些细菌被抗生素杀死,但阻止空洞痊愈的因素目前尚不清楚。
由于空洞中的MTB合成并释放了大量TDM,因此推测TDM可能参与了空洞的维持。在空洞中,MTB几乎完全接近于气液界面生长。这些微生物在气液界面很可能以菌膜形式生长。分枝杆菌的由来就是因为它们倾向于以很厚的菌膜形式生长,类似于霉菌在液体表面生长[29]。这种空洞表面覆盖的膜样物质与MTB的菌膜类似[13]。毒性MTB以菌膜方式生长会释放TDM,从而以非常坚硬的薄层分布于物体表面,并促使MTB以索状形式分布在表面并攀爬上锥形瓶的壁上[47]。由于存在于气液界面的毒性TDM分子单层非常类似于其在油或塑料表面一样(图1B),因此一个开放的空洞具有持续产生毒性和抗原的内表面,从而杀死巨噬细胞,阻止自愈,维持了空洞存在,使MTB不断从空洞排出,并通过咳嗽进入到环境中。因为TDM分子单层比宿主的任何结构都要坚硬和稳定,所以其毒性和抗原性使得具有足够免疫力的宿主空洞能够持续存在。或许这是MTB将患者变为细菌雾化器,为其持续感染新的个体,保证其持续存在的机制。
讨 论
即使在同一个体肺的其他部分具有强烈炎症反应,继发性感染发展的早期阶段也能够在肺泡的某个部分长期无症状聚集分枝杆菌抗原,并表现出很少或没有炎症反应[13]。这就产生一个问题:为什么在肺的一部分存在强烈的过敏性炎症反应,而在其他部分却表现出非常少的炎症反应。除了继发性感染起始于不运动、低通气量和低血流灌注肺尖部位外,Hunter等[33]提出一个假设,认为另一个主要原因是由于细菌表面固定的海藻糖形成了一个“看不见的斗篷”。该假设解释了自MTB被发现以来病理学家一直困惑的问题。
海藻糖可以影响分子甚至整个细菌与水的相互作用。某些植物、昆虫和真菌能够在极端干燥的环境中存活,主要原因是它们具有游离的海藻糖分子,当水蒸发后其能够取代水分子维持蛋白和脂质的构象。当水恢复后,水又能够取代海藻糖,从而使这些生物恢复活性。
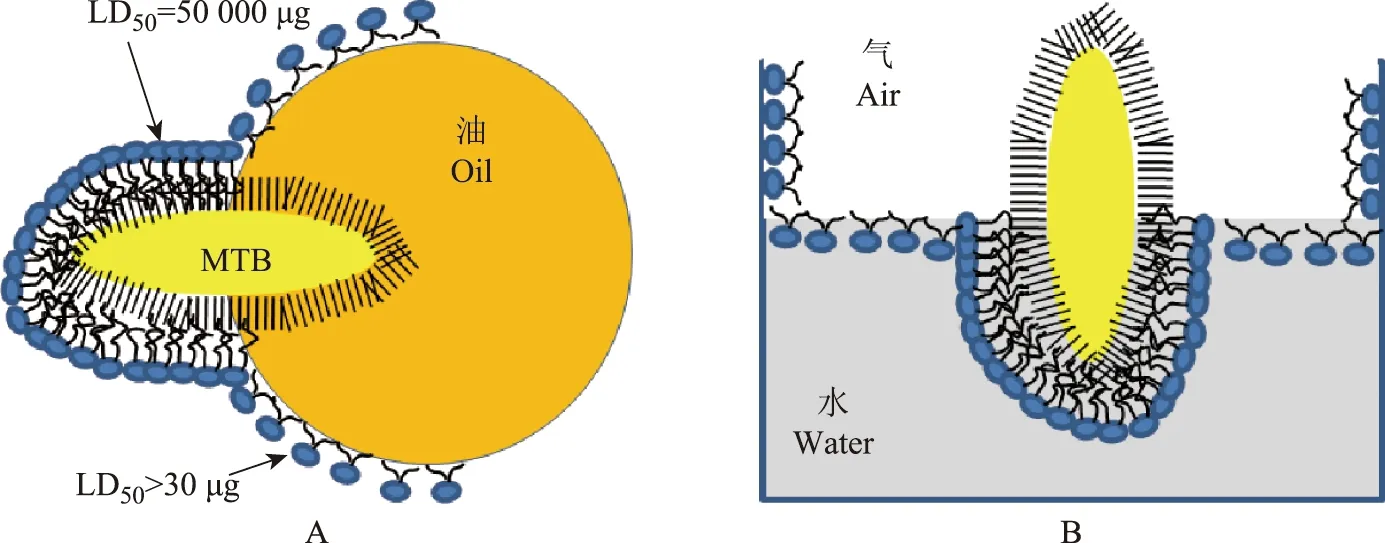
MTB:结核分枝杆菌;TDM:海藻糖-6,6’-二分枝菌酸MTB:mycobacterium tuberculosis;TDM:trehalose-6,6’-dimycolateA.MTB细胞表面的TDM转移到油滴表面的示意图;B.在空气-水界面形成的毒性单分子层示意图A.diagram of MTB transferring TDM to the surface of an oil drop;B.diagram of the formation of the toxic monolayer at an air-water interface图1 MTB细胞表面的TDM的毒性可以通过与脂质和空气相互作用激活Fig 1 Activation of the toxicity of TDM from MTB by interaction with lipids and air

单层分子结构产生于疏水界面,分枝杆菌细胞壁非常厚,主要由脂阿拉伯甘露聚糖和与细胞膜平面垂直排列的分枝菌酸组成,从而与TDM产生了交错双分子层;在水悬液中,TDM形成圆筒状微粒,其表面完全由海藻糖部分组成,脂肪酸部分位于内部The structure of the crystalline monolayer that forms at hydrophobic interface,the mycobacterial cell wall is thick,consisting mainly of lipoarabinomannan and mycolic acids that are oriented perpendicular to the membrane plane to produce an intercalated bilayer with TDM;in aqueous suspension,TDM forms cylindrical micelles whose surface consists entirely of trehalose moieties,with fatty acid moieties inside图2 TDM形成的3种稳定结构Fig 2 Three stable organized structures of cord factor
TDM微粒和双层结构非常稳定,这是由于内部是由极长的分枝菌酸链相互结合,外面是由海藻糖形成坚硬表面的缘故。这些表面的海藻糖分子能够与水结合并降低它们的运动(图3A、B)。任何受体在遇到这样的表面时都只能与表面结合的水接触。因此,分枝杆菌表面包被的TDM以微粒或双分子层存在从而形成了“看不见的斗篷”[33]。MTB的看不见的斗篷使得这些配体“看不见”受体,这种方式导致MTB能够感染很多类型的细胞并持留其中,而在其他地方具有强烈免疫应答、严重的炎症反应和肉芽肿形成。这也是继发性感染中极为重要的一环,可以保证MTB能够无症状的聚集分枝杆菌抗原和宿主脂质成分。这个“看不见的斗篷”可能给MTB提供至少两种操控其周围环境的能力。首先,MTB不必进化出中和或逃避各种细胞攻击的能力;其次,MTB能根据自身需要选择释放信号分子以操控宿主细胞的功能。
在继发性感染早期,肺泡中感染的巨噬细胞可以无症状存在几个月,在此过程中MTB安静地合成分枝杆菌抗原[13]。而这些都发生在具有最小炎症的区域。因此,这个“看不见的斗篷”在早期继发性感染的发展过程中非常有效。该方式导致MTB能够无症状的为突然的大面积坏死反应准备条件,从而产生空洞,为感染新的宿主提供机会。
TDM的单层分子也同样有趣。TDM是到目前为止所认识的MTB毒性最大的脂质成分,但是仅以单层分子形式出现时才具有毒性。当TDM覆盖在颗粒表面并以单层形式出现时,颗粒直径大于25 μm后毒性随颗粒直径增加显著增加[44]。TDM单层分子的物理结构和生物学活性类似于二氧化硅晶体:鳞石英、方石英、石英和超石英[33,36]。这些晶体都是纯的二氧化硅,但在晶体结构上有所不同。超石英是最致密的晶体形式,无毒;鳞石英是最疏松的晶体形式,毒性最强。超石英具有类似于TMD微粒的规则表面,而鳞石英具有类似于TMD单层分子那样规则的亲水和疏水间隔的表面[48]。此外,石英的毒性随着颗粒大小增加显著增加[49]。石英和TDM都能够诱导肉芽肿形成和增加肺结核发生[36]。最后,毒性的二氧化硅晶体类似于TDM单层分子,能够破坏膜结构,杀死与之接触的细胞[29]。
水是维持生命存在的各种分子的溶剂。研究显示水是生物学活动中的积极参与者,尤其是在界面上发生的生物学反应[48]。水分子的氢原子能够与周围其他水分子强烈相互作用从而形成网状晶格。在几何上,这个网格不可能在很大范围的疏水表面上保持完整性。如果相互作用表面对水分子的亲和力远高于水分子之间的亲和力,就会发生快速脱水[49],从而导致表面的“干燥”并形成非常薄的蒸汽层将液体与表面分开[48],这会对水分子产生更大的表面张力。因此,附近任何能够释放这种张力的分子或颗粒被迫留在表面以减轻这种张力,从而产生一种黏附作用,而这种黏附作用随着水分子的取代而消失。然而,靠近含水的表面会导致分子或颗粒的侧向运动,离开疏水区域,并被其他分子取代。因此,这种方式或许是TDM的毒性机制。水分子氢键的断裂对水分子产生了更大的张力,导致其向附近的膜或蛋白移动[50](图3C)。在毒性晶体结构表面上交替存在的亲水和疏水结构区域产生了很强的黏附作用,缓解了对水的张力和降低了侧向运动,导致细胞在各个方向的快速伸展,最终造成细胞膜的破裂。这类似于很多催化剂的作用机制,其需要紧密黏附以促进化学反应,以及随后产物的侧向运动,从而反应可以重复进行。
圆筒状微粒和交错双层分子的表面完全是海藻糖;海藻糖分子与水结合,外表覆盖上水分子后阻止任何受体与其结合;TDM单层分子扭曲了周围的水分子,虽然水分子能够与海藻糖结合,但不能与暴露在外的脂肪酸结合,而这种交错的分子结构非常稳固足以对抗水的作用力,从而产生结合和破坏与之接触的细胞膜Both the cylindrical micelle and intercalated bilayer have surfaces that are radically immobilizedtrehalose;trehalose binds with water and the surface is coated with water molecules to prevent any receptors from binding to it;TDM monolayer distorts surrounding water:the water molecules can bind with trehalose while not with the exposed fatty acids;the interlocking molecular structure of mycolic acid chains is stable enough to withstand the force of water,resulting in the binding and the destruction of the cell membrane that makes contact图3 不同形式的TDM对周围水分子的影响Fig 3 Influences of different TDM configurations on surrounding water molecules
MTB具有逃避宿主防御的独特能力,存活于人类组织中,最终逃出宿主并感染新的宿主。因为MTB是专性人类病原菌[51],所有基因都被选择以更适应在人类中存活和传播。由于MTB需要消耗大量能源合成TDM,并将几乎纯的TDM武装到细胞表面,因此TDM对其生存至关重要。通过TDM,MTB能够调控其周围环境的生物物理性质。对这些机制的进一步研究将为了解MTB独特的致病机制提供新的认识。
综上,MTB是人类专性病原菌,因为只有人类会产生原发性感染和继发性感染,并通过继发性感染传播到新的宿主[51]。原发性感染可通过产生有效的系统免疫保护宿主不会因为感染而死亡,为病原菌提供长期持留的宿主;而继发性感染产生的空洞为MTB进入环境传播到新的宿主提供机会。由于缺乏有效的模型,结核病发病机制的许多基本问题仍然是现代科学无法解决的。本文提供了一种可能的继发性感染发展机制,阐述了继发性感染如何分离出一部分肺以促进宿主脂质成分和分枝杆菌抗原的聚集,为产生干酪样肺炎和空洞的超敏反应创造条件。脂质成分的聚集导致脂质微滴的大小逐渐增加,当包被有TDM的脂质微滴大小增加到临界值时导致TDM构象改变、毒性增强,并诱导大量肿瘤坏死因子释放,触发突然的干酪样肺炎,导致空洞产生。在空洞中MTB以生物膜的形式生长,生物膜方式存在的MTB不仅与组织的坏死、空洞形成相关,而且还涉及感染的发生以及结核病的复发[52-53]。
