利用下颌骨形态鉴别野生和饲养北美水貂的研究
2021-05-06刘博洋孟庆博祁永来徐艳春杨淑慧
刘博洋 孟庆博 祁永来 徐艳春,2,3 华 彦 杨淑慧*
(1.东北林业大学野生动物与自然保护地学院,哈尔滨,150040;2.国家林业和草原局野生动植物检测中心,哈尔滨,150040;3.国家林业和草原局野生动物保护与利用工程技术研究中心,哈尔滨,150040;4.广东省森林培育与保护利用重点实验室,广州,510520)
由于过度利用、栖息地丧失、人类干扰等多种原因,野生动物的生存一直受到威胁,种群数量大幅度减少,很多物种已无法作为资源使用。随着人工养殖产业的不断扩大,野生动物产品的相关贸易市场被放大,在经济利益的驱使下,一些投机分子将野外盗捕的动物假借人工饲养的名义进行贩卖。如果无法正确区分其来源,执法者就无法甄别违法犯罪行为,盗猎的动物就会在合法养殖业中被洗白,从而使养殖业成为盗猎活动的保护伞。所以,鉴别动物来源于养殖场还是野外是保护野生资源,维护养殖产业正常秩序的关键。
动物头骨的结构复杂,常表现明显的适应性变化[1-2]。如因营养生态位的分离而产生的性别差异[3-4]、不同方向的自然选择产生的地理种群间、亚种间和种间的变异[5-9]。甚至于在不同环境下头骨的发育都有着明显的差异[10-11]。骨骼由胶原纤维按照一定的方式排列、编织,矿物质沉积在胶原纤维上,骨骼的应力在骨骼发育过程中,会影响胶原纤维的编织方式[12]。动物捕食和进食过程中需要撕裂、切割和咀嚼等动作,但这些动作的使用频率和强度因食物的质地而异,进而影响头骨的形态发育。研究显示,野生动物获取食物的坚硬程度不同时鼻口部宽度、颂骨的宽度会发生变化[13],同时形状[14]、冠状突长度及枕骨高度也会变化[15]。此外,骨骼是由蛋白质和矿物质等基本原料构成的。骨骼发育过程中这些基本物质缺乏或不平衡,会直接影响骨骼的结构和强度,进而产生一定的偏离正常状态的形变。如食物中蛋白质缺乏会影响头骨骨头的生长速度[16-18]。
人工饲养的野生动物,尤其是商业化饲养的动物,所处空间较小,食物通常以人工配合饲料为主,质地柔软,饲料原料种类较少,营养成分稳定,而野外动物食谱广泛,质地差异大,动物所获营养成分依食物种类而变化。我们推测,动物在采食过程中,撕咬、咀嚼、切割的力量、频度和持久性均有差别,这些差异通过改变应力来影响头骨的形态,尤其是下颌骨的形态,进而为鉴别野生和人工养殖个体的提供可能。
食肉动物的臼齿和前臼齿是切割食物的主要工具,上颌的齿尖与下颌的凹槽相对,下颌的齿尖与上颌的凹槽相对,从而形成锯齿剪刀一样的结构,既能固定食物的位置,又能加大压强和剪切力。可以推想,动物生长过程中,下颌骨因为食物质地的不同需要不同的咬合力,野外动物所需的咬合力大,而人工饲养动物所需的咬合力较小。由此形成一个剪刀状的杠杆系统:下颌骨的髁状突为支点,以咬合肌群为动力,以牙齿的齿尖为阻力点,以髁状突到齿尖的距离为力臂。要提高咬合的效率,缩短力臂是最好的策略。这种策略可能在个体发育过程中体现出来:食物柔软的动物咬合力臂可能较食物坚硬的动物长些。如果这一推想是正确的,则咬合杠杆系统的结构特点就能够用来鉴别野生和饲养的食肉动物。
本研究以人工饲养和野外的北美水貂(Neovisonvison)为代表,定量比较下颌骨的形态在两组之间的差异,并据此建立判别来源的指标。
1 材料与方法
1.1 头骨样品及其预处理
本研究的实验动物为野外与人工饲养的北美水貂。北美水貂在19世纪80年代末在加拿大笼养成功,20世纪30年代率先在欧洲建立养殖场,50年代引进我国进行规模化饲养。从30年代正式农场饲养至今已经繁育80余代。目前我国野外的水貂均为养殖场逃逸个体所建立的种群。本研究采用的野生水貂25只,均来自大兴安岭北部的漠河县,在2011年11月—2012年1月从野外捕获,包括雄性个体7只,雌性个体18只;人工饲养的水貂38只,来自大兴安岭根河市阿龙山镇的水貂饲养场。
从颈椎处切下头颅,做好标记,纱布包裹后放入沸水中煮至肌肉变软,用手术刀去除头骨上附着的肌肉、结缔组织,放入含有0.1%的Na2CO3的温水中去除油脂,取出经清水漂洗后自然干燥备用[19]。
1.2 下颌骨及牙齿各项量度的测量方法
首先测定2组水貂下颌骨的量度指标,如图1所示。
L1:下原尖(protoconid)到髁状突的距离。
L2:下前尖(paraconid)到髁状突的距离。
L3:前臼齿第3齿尖到髁状突的距离。
L4:犬齿齿尖到髁状突的距离。
L:髁状突到下颌骨最前端距离,即下颌长。
L′:髁状突到犬齿唇面基部与下颌骨结合处的距离。
M1:臼齿长度。
M2:前臼齿第3齿基部长度。
H1:臼齿下原尖的高度,即下原尖顶端到其牙冠与下颌骨接合处的垂直距离。
H2:臼齿下前尖的高度,即下前尖顶端到其与下颌骨接合处的垂直距离。
H3:前臼齿第3齿尖顶端到与下颌骨接合处的垂直距离。
H4:犬齿高犬齿尖端到其与下颌骨接合处的距离。
分别将犬齿尖端,犬齿齿冠与下颌骨交点,髁状突记标记点,作为三角形的3个顶点,该三角形上个边长分别为L4、H4及L′,将H4与L′所组成的角记为∠A,由余弦定理计算CosA:
为了消除个体大小的影响,用上述各项量度定义了13个判别指标,用以判别来源,它们分别是:
L1/L:以下颌为标准,下原尖力臂的相对长度。
L2/L:以下颌为标准,下前尖力臂的相对长度。
L3/L:以下颌为标准,前臼齿第3齿尖力臂的相对长度。
L1/M1:以齿尖所在齿齿长为标准,下原尖力臂的相对长度。
L2/M2:以齿尖所在齿齿长为标准,下前尖力臂的相对长度。
L3/M3:以齿尖所在齿齿长为标准,前臼齿第3齿尖力臂的相对长度。
H1/L1:以下原尖力臂长为标准,下原尖高的相对长度。
H2/L2:以下前尖力臂长为标准,下前尖高的相对长度。
H3/L3:以前臼齿第3齿尖力臂长为标准,前臼齿第3齿尖高的相对长度。
H1·L1/L2:下原尖的相对发育情况。
H2·L2/L2:下前尖的相对发育情况。
H3·L3/L2:前臼齿第3齿尖的相对发育情况。
H4·L4/L2:犬齿的相对发育情况。
1.3 数据处理
人工饲养水貂与野外水貂各项指标差异的检验使用独立样本t-检验(α=0.05)。各项量度或指标与L或L2的回归分析中,首先对数据进行对数转换,然后采用一元线性回归分析,计算R2值,并检验回归的显著性(α=0.05)。回归关系用散点图显示。对各项指标在来源或性别判别中的有效性评估采用判别分析法,以已知样本的正确回判率(%)为评估指标,指标越接近100%有效性越高。所有的统计分析采用SPSS Statistics 19.0(IBM)完成。
2 结果
2.1 各项量度性别二型性
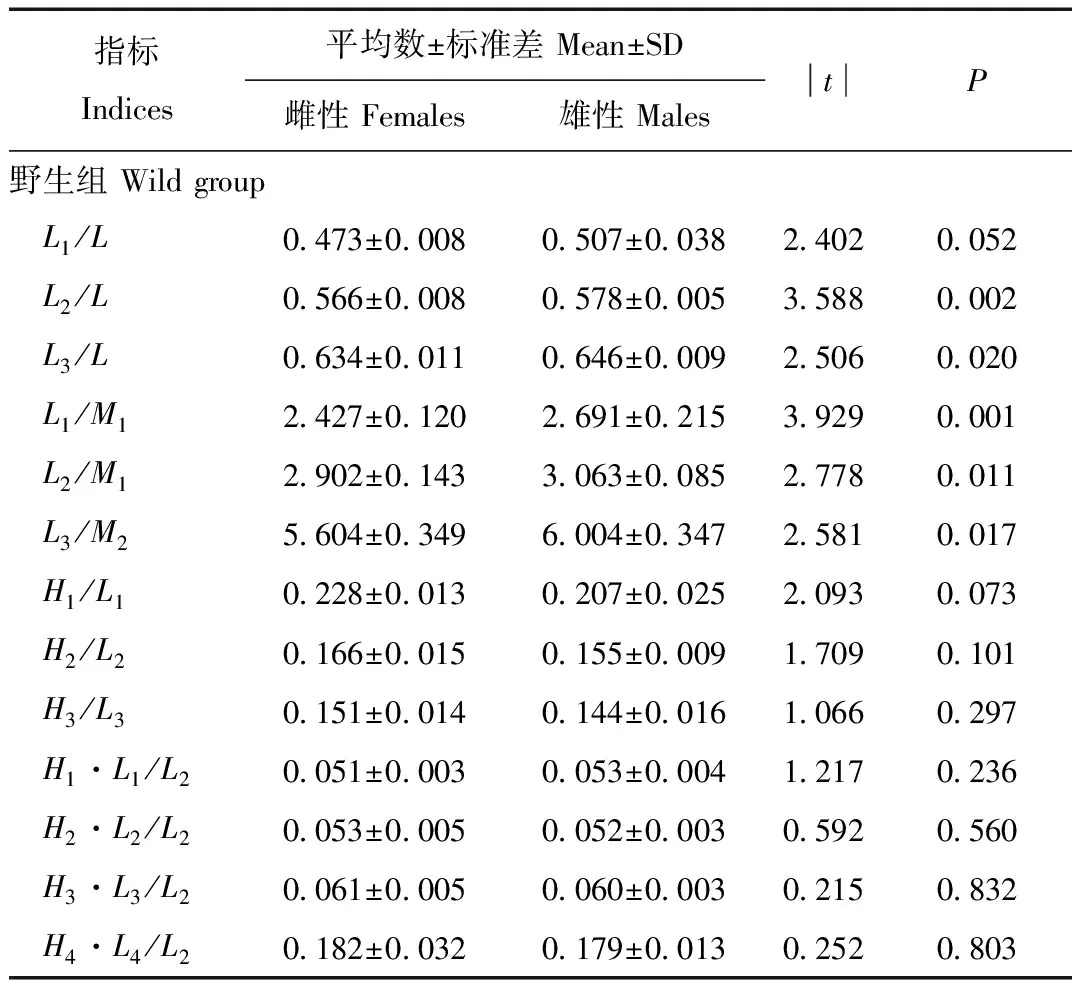
如表1、表2所示,力臂长度指标L1、L2、L3、L4、L,臼齿长度指标M1、M2,齿尖高度指标H1、H2、H3、H4等均存在显著的二型性。独立样本t检验显示,饲养组中,这些指标在性别间差异显著(P<0.001),且均为雄性显著大于雌性。野生组中,这些指标也显示出雄性显著大于雌性的趋势(0.001 表1 野生和人工饲养的北美水貂下颌骨的量度 续表1 表2 野生和人工饲养北美水貂下颌骨形态指标的差异 如表1所示,L1、L2、L3、L4、L、M1、M2、H1、H2、H3、H4在饲养组的平均值均显著大于野生组(P<0.001)。CosA在饲养组和野生组之间存在显著差异(t=-5.183,P<0.001),且野生组中明显大于饲养组。余弦函数在0—180°间为减函数,所以饲养组个体中的∠A较野生组个体大。 这些指标在野生组和饲养组之间的显著差异说明它们能够提供鉴别水貂来源的相关信息。但是这些指标受到身体比例关系的影响,不能直接用于判别,所以,我们建立了13个相对判别指标。 为了在回归分析中,数据符合或近似于正态分布,将各项量度指标进行了对数转换,即取其常用对数值作为变量,然后进行比例分析。 以下颌骨长的对数值LogL为标准,采用线性回归分别分析LogL1、LogL2、LogL3、LogM1、LogM2、LogH1、LogH2、LogH3与其比例关系。结果如图2,LogL1、LogL2、LogL3、LogM1、LogM2、LogH3等与LogL均存在显著的线性关系(野生组0.001 以LogL2为标准,对LogL1·H1、LogL2·H2、LogL3·H3、LogL4·H4进行回归分析,结果如图3,LogL1·H1、LogL2·H2、LogL3·H3、LogL4·H4等与LogL2均存在显著的线性关系(野生组P<0.001,饲养组P<0.001)。 由于存在性别二型性,性别可能影响来源判别的正确率。因此,对所有判别指标进行筛选,挑选出在野生组和饲养组中不存在二型性的判别指标用于判别分析。如表3,在两组中均不存在二型性的判别指标包括:H2/L2(野生组t=1.709,P=0.101;饲养组t=0.902,P=0.373)、H3/L3(野生组t=1.066,P=0.297;饲养组t=1.095,P=0.281)、H1·L1/L2(野生组t=1.217,P=0.236;饲养组t=0.722,P=0.475)、H2·L2/L2(野生组t=0.592,P=0.560;饲养组t=0.064,P=0.949)、H3·L3/L2(野生组t=0.215,P=0.832;饲养组t=0.519,P=0.607)、H4·L4/L2(野生组t=0.252,P=0.803;饲养组t=1.766,P=0.087)、CosA(野生组t=1.074,P=0.294;饲养组t=1.372,P=0.179)。 表3 野生组、饲养组性别二型性的分析 续表3 对表3中筛选出的无二型性的判别指标进行差异分析,如表4,H2/L2(t=5.727,P= 0.000)、H3/L3(t=3.195,P=0.002)、H1·L1/L2(t=2.446,P=0.019)、H2·L2/L2(t=5.132,P=0.000)、H3·L3/L2(t=3.128,P=0.003)、CosA(t=5.183,P=0.000)在野生、饲养组间存在显著差异,H4·L4/L2(t=1.813,P=0.075)在野生、饲养组间无显著差异。对在两组间表现出显著差异的判别指标进行来源有效性分析,结果如表5所示:判别指标H2/L2+CosA对已知来源个体的正确回判率为91.2%,其余各项判别指标正确回判率为60.3%—73.3%。 表4 无二型性判别指标的组间差异分析 表5 各项判别指标对来源判别的有效性 本研究采用的北美水貂均源自人工驯养的个体,其中一些个体逃逸到野外并建立了种群[20]。野外的水貂和养殖场的水貂从遗传背景上比较相近,但是在头骨的形态上却表现出较大的差异。仅从下颌骨来看,下颌长度、牙齿高度、宽度、犬齿角度等都在两组之间发生显著差异(表2)。食肉动物从幼年到成年自行捕食的不同阶段,臼齿、犬齿利用的强度和模式不同,下颌骨、下颌骨的愈合带和牙齿的力学结构都会产生相应的变化[21]。雌雄赤狐(Vulpesvulpes)幼年时食性的差异(种类和质地)与其头骨形态二型性的形成密切相关,雄性较多进食绵羊尸体,头骨更加强壮,咬合力更大,而雌性食谱以啮齿类、无脊椎动物为主,则其咬合力较低,头骨也不够粗壮[22]。模式动物研究进一步表明,咬合力对臼齿齿根的长宽均有影响,增加咬合力可使齿根从狭长变得短粗[23]。这些结果预示着人工饲养环境和野外环境下,捕食、防卫等行为对下颌骨的要求不同是导致水貂下颌骨形态差异的重要原因。 我们发现臼齿下原尖和下前尖的相对力臂长在野生组均显著小于饲养组。这意味着,相比饲养组,野生个体的下原尖、下前尖更靠近髁状突。这样,在相同的咬合力的情况下,野生水貂的下原尖和下前尖上产生更大的压强,对食物的切割更为有效。此外,相对于下颌长,臼齿齿冠相对长度、齿尖相对高度在野生组也显著大于饲养组。这说明野生个体有更长的臼齿齿冠,使切割食物时,臼齿与食物接触面更大,有利于切割较大的食物。综合来看,本研究的结果支持了我们的推测,即野生水貂较人工饲养的水貂在切割食物的效率更高。 犬齿与下颌骨夹角的余弦值CosA在野生组也显著大于饲养组(P<0.001)。余弦函数在0—180°间为减函数,因此,饲养组个体犬齿与颌骨夹角大于野生组个体,也就是说,饲养组水貂的犬齿更加向前伸展。这种差异可能来自饲养水貂的刻板行为。水貂被饲养在狭小笼舍中,环境缺乏变化,饮食也较为固定,容易引发刻板行为[24],其中无目的的撕咬笼舍以及其他物体是大多数养殖场都存在的刻板行为之一。用犬齿撕咬物体对犬齿产生向外的拉力,在个体发育过程中,这种拉力可以改变犬齿原有的角度,变得向外倾斜。向前倾斜会降低撕咬的效能。 在下颌骨形态的众多变化中(表2,表3,表4),在野生和饲养组之间的差别最大的还数臼齿下前尖处的齿高与其力臂的之比,以及下颌犬齿向前伸展的角度(∠A,图1)两个指标,而且它们没有性别二型性(表5)。利用H2/L2+CosA对已知来源个体进行回判,正确回判率达到91.2%,远优于其余各项判别指标(正确回判率为60.3%—73.3%)(表5)。因此,这说明,下颌骨和犬齿的形态可以有效判别野生和人工饲养的水貂。那么,水貂作为典型的小型食肉动物,在生态习性、行为模式等方面具备其他中小型鼬科(Mustelidae)动物的典型特征,所得到的结果应该具有一定的推广意义,值得在其他物种进行尝试。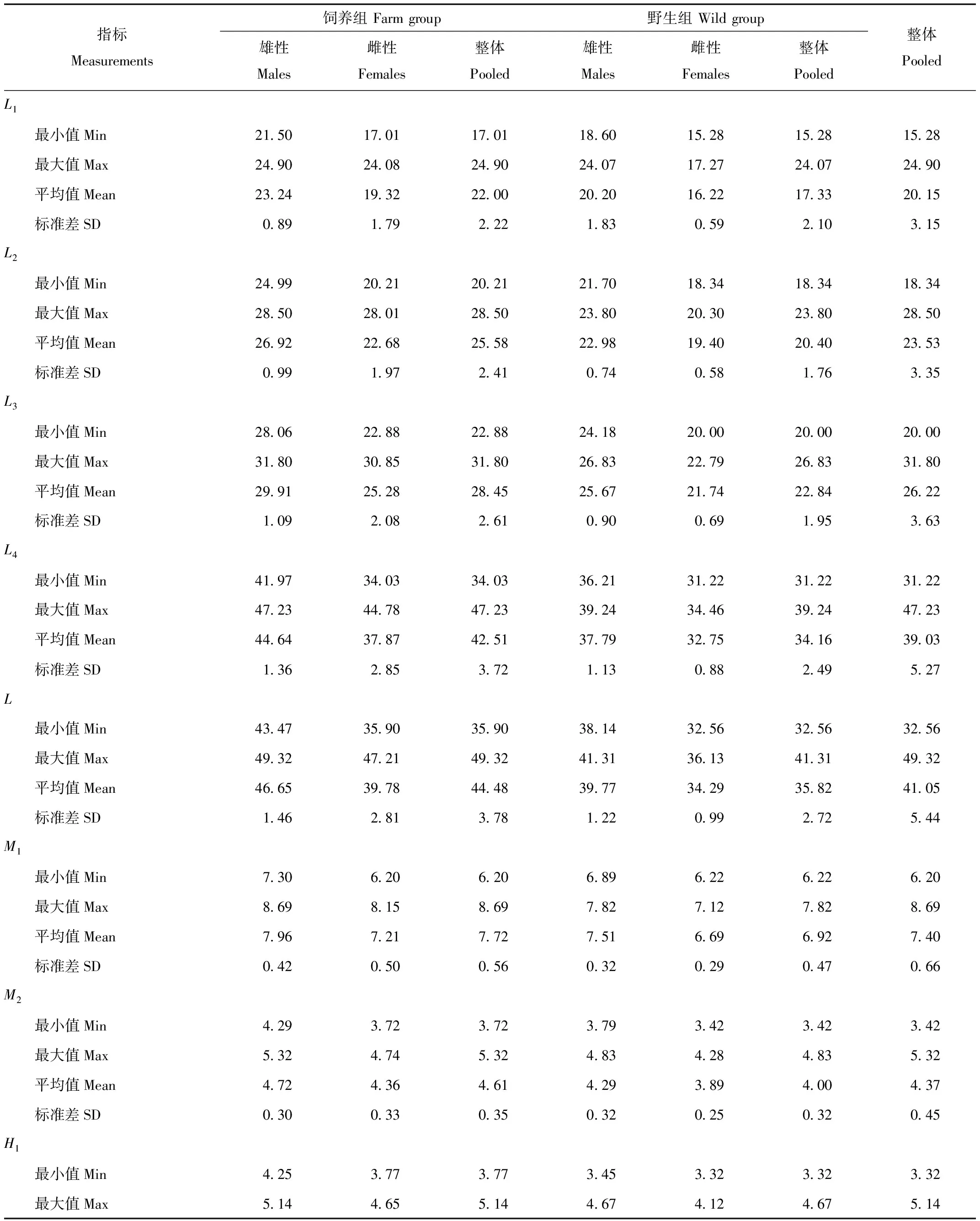


2.2 各项量度指标在野生组和饲养组之中间的差异
2.3 各项指标间的比例分析
2.4 各项判别指标性别二型性

2.5 各项指标来源判别有效性的评估


3 讨论
