乌司他丁对大鼠脑缺血再灌注后海马NMDAR1 表达的影响
2021-04-28董真真
董真真 袁 峰 白 倩
郑州大学第二附属医院,河南 郑州450003
脑缺血疾病是临床一种常见病、多发病,主要有效治疗方法为早期恢复缺血半暗带血供,但血管再通后其功能尚不能恢复,却出现脑功能损害加重的情况即为脑缺血再灌注损伤。脑灌注损伤是决定疾病预后的重要因素,因此如何最大程度降低其损害,提高神经元的存活率成为研究脑灌注损伤的主要课题。脑缺血再灌注损伤是一种多机制、多因素介导的细胞凋亡、神经受损过程。谷氨酸、门冬氨酸等兴奋性氨基酸是中枢神经系统重要的兴奋性神经递质。研究表明,兴奋性氨基酸在脑缺血再灌注后释放增加,是脑缺血再灌注损伤的重要机制[1],对脑组织的损害作用是通过作用于相应的受体而产生。谷氨酸受体包括离子型受体和代谢型受体,离子型受体直接激活受体使离子通道开放,代谢型受体通过激活G 蛋白和第二信使间接使离子通道开放,其中N-甲基-D-天门冬氨酸受体(NMDAR)是最重要的谷氨酸离子型受体,兴奋性氨基酸通过作用于NMDAR 使大量Ca2+内流,致细胞内Ca2+超载,进而引起一系列病理变化致神经细胞死亡。研究表明,NMDAR 参与兴奋性氨基酸的毒性作用,在局灶脑缺血再灌注后表达增加[2]。因此,抑制NMDAR 的表达有可能降低脑缺血再灌注损伤。NMDAR 抑制剂分为竞争性拮抗剂和非竞争性拮抗剂,如地卓西平和氯胺酮被证实能减轻缺血后脑损伤[3-4],但同时会引起神经元空泡形成,造成行为和认知等精神症状的不良反应[5]。NMDAR 有3 种亚基:NMDAR1、NMDAR2、NMDAR3,其中NMDAR1 是Ca2+通道的调节者,是介导谷氨酸兴奋毒性的主要组分[6]。
乌司他丁是一种能抑制多种蛋白水解酶活力的蛋白酶抑制剂,常用于急性胰腺炎、循环衰竭的救治。研究发现,乌司他丁能降低围术期应激反应,抑制炎症介质的释放,从而保护心、肝、肾等重要脏器功能[7];也可减轻小鼠脑缺血再灌注损伤[8],用于缺血再灌注损伤能够减少氧自由基的释放和Ca2+内流,降低Ca2+超载及兴奋性氨基酸的释放[9]。本研究拟通过探讨乌司他丁对大鼠脑缺血再灌注时海马NMDAR1表达的影响评价其神经保护作用。
1 材料和方法
1.1 材料
1.1.1 动物与分组:清洁健康雄性SD 大鼠72 只(河南实验动物中心),体质量220~280 g,将大鼠随机分为假手术组(S组)、脑缺血再灌注组(I/R组)、乌司他丁干预组(U组),每组24只。
1.1.2 主要仪器和试剂:RT-PCR 图像分析系统(赛沛GeneXpert Infinity);PCR 仪(美国,Applied Biosystems),电泳仪(北京市六一仪器厂,DYY-11),图像采集系统(VisionBank SVS),乌司他丁(广东天普生化医药股份有限公司,国药准字号H19990133),TUNEL 试剂盒(罗氏ZK 8005),PCR 试剂盒(DRR036A PrimeScript,大连宝生物工程有限公司),兔抗大鼠NMDAR1 多克隆抗体(Sigma 公司,美国),免疫组化试剂盒(SABC 试剂盒,武汉博士德生物工程有限公司),DAB显色试剂盒(上海康朗生物技术有限公司)。
1.2 方法
1.2.1 脑缺血再灌注模型的制备:参照LONGA等[10]线栓法制备大鼠右侧大脑中动脉缺血模型。禁食、禁水12 h,用10%水合氯醛3.5 mL/kg 进行腹腔注射麻醉,将大鼠放在操作台上,仰卧固定四肢,取颈部正中切口,剪开皮肤、钝性分离组织肌肉,暴露颈总动脉、颈内动脉、颈外动脉,结扎颈外动脉,将线栓经颈总动脉插入颈内动脉约20 mm 固定,清创缝合伤口。缺血2 h后再缓慢拔出栓子,使栓子头端回到颈内动脉起始部,恢复组织灌注。S组仅暴露右侧颈总动脉后缝合,不做其余处理。U组于再灌注即刻腹腔注射注射乌司他丁20 000 U/kg,S 组及I/R 组用相同剂量的生理盐水替代。
1.2.2 神经功能评分(NDS):参照LONGA[4]的评分方法,再灌注3 h 后观察并记录神经功能缺失症状;无神经损伤征象为0分,不能完全伸展健侧前肢为1分,行走时向健侧旋转征象为2 分,向健侧跌倒为3分,无自发活动及意识障碍者为4分,1~3分为模型制作成功。记录各组大鼠神经功能缺陷评分。
1.2.3 凋亡细胞数的检测:NDS完成后,每组随机抽取8 只大鼠,4%多聚甲醛灌注固定后,快速断头取脑,取病变侧视交叉后2~4 mm做冠状切片,连续制成厚约5 μm 的切片。按照TUNEL 法试剂盒步骤操作,胞核染成棕褐色即为阳性细胞。将每只大鼠均匀切片,随机选择不连续的切片5 张,每张切片的5个不同视野在400倍镜下进行观察并计数,随后求其平均数。
1.2.4 RT-PCR 检测海马神经元NMDAR1 mRNA 水平的表达:NDS完成后,每组随机抽取8只大鼠,迅速断头取脑,将脑组织研磨提取RNA,按照TaKaRa RNA PCR试剂盒步骤进行。以cDNA模板行PCR扩增:NMDAR1(512 bp)上游引物:5’-GACTATACCAGCCGAATAAGCG-3’,下游引物:5’-CTGCTACAGCAGACTCGTCACAT-3’。加入适量的ddH2O,使体积增大,轻轻混匀、离心。设定PCR 程序,在适当的温度扩增30个循环,于PCR仪中获取所需要的产物,行琼脂糖凝胶电泳,采用凝胶成像分析仪对结果进行观察、拍照,并测定每条带的积分光密度值。
1.2.5 采用免疫组化法检测NMDAR1 的蛋白表达:NDS 完成后,各组取8 只大鼠,取视交叉神经起点处前后各2 mm 的冠状切片,放入4%多聚甲醛中4 ℃固定18 h,经15%、20%和30%的蔗糖梯度脱水后在Leica 冰冻切片机上连续切片,片厚15 μm。免疫组化染色:切片后依次用4%多聚甲醛固定40 min,滴加内源性过氧化物酶阻断剂、山羊血清各40min,滴加稀释的兔抗NMDAR1 多克隆抗体(1∶500)浸泡于4 ℃的湿盒过夜,洗膜后加入链酶亲和素过氧化物酶37 ℃30 min 进行二抗反应;然后加入新鲜配制的DAB 显色液镜下观察5 min,用1%苏木素复染,蒸馏水冲洗、脱水,中性树胶封片,光镜下观察观察缺血侧海马CA1区NMDAR1的表达,用计算机自动图像分析软件计算缺血再灌注侧海马区NMDAR1 表达的平均光密度值(mean optical density,MOD)。
1.3 统计学方法 采用SPSS 20.0 统计软件进行统计学分析,正态分布的计量资料用均数±标准差(±s)表示,采用t检验,组间比较采用单因素方差分析,以P<0.05为差异有统计学意义。
2 结果
2.1 乌司他丁对大鼠神经功能和脑梗死体积组织凋亡细胞数的影响 与S组比较,I/R 组和U组NDS评分和脑组织凋亡细胞数增加(P<0.05);与I/R 组比较,U 组NDS 评分和脑组织凋亡细胞数降低(P<0.05)。见表1。
2.2 乌司他丁对缺血再灌注大鼠海马CA1 区NMDR1 mRNA 水平表达和NMDAR1 蛋白表达的影响 与S 组比较,I/R 组NMDAR1 mRNA 表达上调和NMDAR1 表达的MOD 值升高(P<0.05);与I/R 组比较,U 组NMDAR1 mRNA 表达下降和NMDAR1 表达的MOD值降低(P<0.05)。见图1~2、表2。
表1 3组大鼠NDS评分和脑组织凋亡细胞数比较 (±s)Table 1 Comparison of NDS and the number of apoptotic cells in three groups of rats (±s)
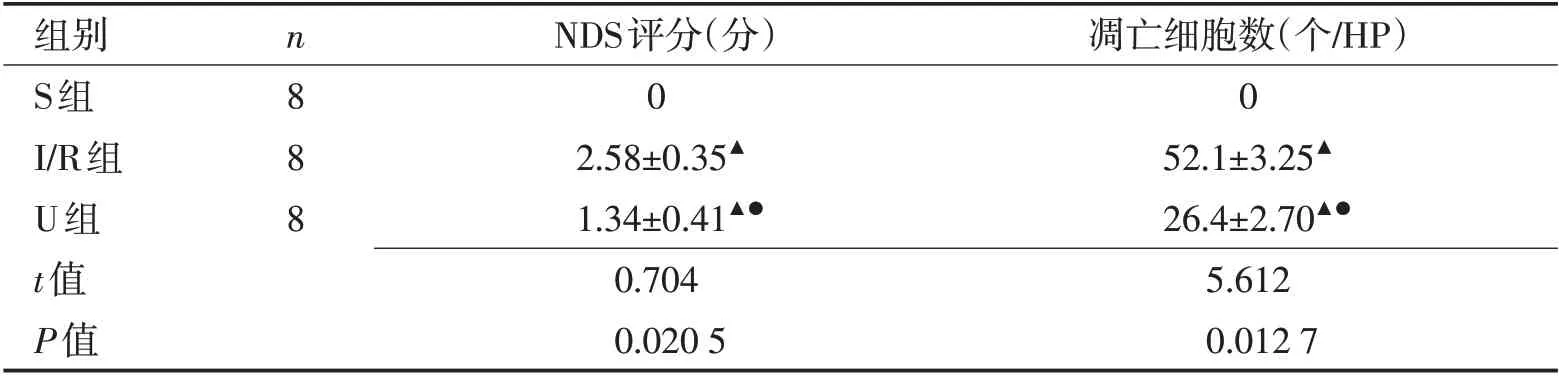
表1 3组大鼠NDS评分和脑组织凋亡细胞数比较 (±s)Table 1 Comparison of NDS and the number of apoptotic cells in three groups of rats (±s)
注:与S组比较,▲P<0.05;与I/R组比较,●P<0.05
组别S组I/R组U组t值P值凋亡细胞数(个/HP)0 52.1±3.25▲26.4±2.70▲●5.612 0.012 7 n888 NDS评分(分)0 2.58±0.35▲1.34±0.41▲●0.704 0.020 5
表2 3组大鼠海马CA1区NMDR1 mRNA水平和NMDAR1表达MOD值比较 (±s)Table 2 Comparison of the expression of NMDR1 mRNA and the MOD value of NMDAR1 expression in the hippocampal CA1 region of the three groups of rats (±s)

表2 3组大鼠海马CA1区NMDR1 mRNA水平和NMDAR1表达MOD值比较 (±s)Table 2 Comparison of the expression of NMDR1 mRNA and the MOD value of NMDAR1 expression in the hippocampal CA1 region of the three groups of rats (±s)
注:与S组比较,★P<0.05;与I/R组比较,■P<0.05
组别S组I/R组U组t值P值MOD值0.006 6±0.000 7 0.059 4±0.011 1★0.006 8±0.000 4★■3.241 0.021 7 n888 NMDR1 mRNA 0.890 2±0.013 6 1.039 8±0.021 1★0.907 3±0.014 2★■0.437 0.046 1
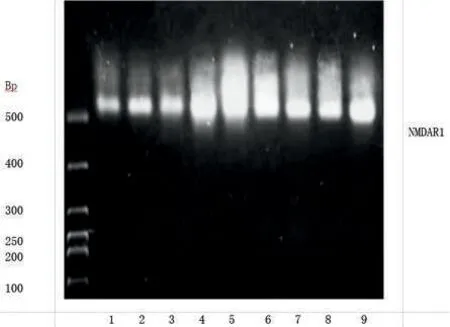
图1 各组大鼠海马组织中NMDAR1 mRNA的表达Figure 1 Expression of NMDAR1 mRNA in the hippocampus of rats in each group
3 讨论
脑缺血再灌注损伤是一种多因素导致细胞发生肿胀、破裂、凋亡的复杂病理过程[11],其机制可能包括兴奋性氨基酸的毒性作用、氧自由基释放、线粒体功能障碍、炎症介质生成、Ca2+超载以及血管通透性改变等[12]。兴奋性氨基酸是中枢系统正常的兴奋性递质,包括谷氨酸和天冬氨酸,其中谷氨酸是神经系统主要的兴奋性递质,参与突触兴奋传导,影响脑功能包括认知、和学习记忆功能[13]。正常情况下,兴奋性氨基酸的产生、摄取和重吸收维持着动态平衡状态。当缺血、缺氧或创伤等损害因素下能促其过度释放产生毒性作用,损伤脑组织[14],其机制为大量兴奋性氨基酸释放引起Na+、Cl-内流、K+外流,导致细胞水肿、变性,蛋白质降解,从而造成不可逆的线粒体坏死和细胞死亡[15-16]。

图2 3组海马CA1区NMDAR1的表达Figure 2 Expression of NMDAR1 in CA1 region of hippocampus in three groups
兴奋性氨基酸对神经细胞产生的毒性作用主要通过相应的受体产生。NMDAR 是一种门控型离子型受体,对Mg2+有电压依赖的阻断作用,有谷氨酸和甘氨酸的结合位点,对Ca2+、Na+、K+均有通透性,因此是最重要的兴奋性氨基酸受体。NMDAR 广泛存在于中枢神经系统,其中以海马、大脑皮质和小脑最为丰富[17]。研究表明,脑缺血再灌注后NMDAR的毒性作用可致记忆细胞发生坏死,也可发生细胞的延迟死亡[18]。研究发现,脑缺血再灌注后损伤后,NMDAR1 mRNA 含量增多和NMDAR1 表达增加[19]。既往将NMDAR 拮抗剂应用于动物模型中研究其抗缺血、缺氧、预防神经元变性的作用,但发现该拮抗剂可损伤呼吸和心血管系统或引起精神系列症状[5],限制了其临床应用。目前已证实NMDAR 有3 种亚基:NMDAR1、NMDAR2、NMDAR3,分别有不同的分布和功能。其中NMDAR1 是Ca2+通道的调节者[20],在脑缺血再灌注中与过量的谷氨酸结合,神经元去极化,大量的Ca2+通道开放,致细胞内Ca2+超载,线粒体膜损伤,能量生成障碍,产生白三烯、前列腺素等炎性介质,进而引起细胞死亡[21]。研究发现,右美托咪定预处理可减少缺血再灌注损伤时兴奋性氨基酸的释放和NMDA受体亚单位NRl的表达而产生脑保护作用[22],布美他尼降低缺血再灌注损伤可能与降低NMDAR1的表达有关[23]。本研究通过检测转录水平NMDAR mRNA 和翻译水平NMDAR 蛋白的表达可以评估对神经细胞的保护作用。研究发现,脑皮质NMDAR1 mRNA 在短暂大脑缺血后再灌注24 h显著增加[24]。也有研究表明,NMDAR1 表达在缺血再灌注3 h 达高峰,然后逐渐下降,12~24 h 后恢复正常[25]。因此,本研究选择灌注3 h 作为观察点,结果显示I/R 组NMDAR mRNA 和NMDAR 蛋白的表达上调,与以往研究结果一致。
乌司他丁是从新鲜人尿中提取的一种能抑制多种蛋白水解酶的蛋白酶抑制剂。研究表明乌司他丁能缓解围术期应激,抑制炎症因子和自由基的释放,对心脏、肝脏等重要脏器的缺血再灌注损伤具有保护作用[26],对中枢的保护作用也已得到证实[27],但其保护机制尚未完全阐明[28]。LI等[29]通过研究乌司他丁对代谢型谷氨酸受体Ⅱ表达的影响评价其脑保护作用;LI等[8]发现乌司他丁通过抑制炎症介质的生成发挥脑保护作用。本研究参照文献[1]制备脑缺血再灌注模型,结果显示I/R组NDS评分和细胞凋亡数增加,说明模型制备成功。U组在灌注即刻给予乌司他丁,结果显示U 组较I/R 组NDS 和细胞凋亡数下降,说明减轻了脑组织的损伤,与以往研究结果相同。U组与I/R组比较,NMDAR1 mRNA表达下降和NMDAR1蛋白表达的MOD值降低,说明乌司他丁下调了NMDAR1 蛋白的表达,可能机制为谷氨酸等兴奋性氨基酸通过受体起作用,受体的活性或数目影响其效能,NMDAR作为谷氨酸的主要受体可能被乌司他丁所抑制;乌司他丁有清除氧自由基和抑制炎症介质释放的作用[30],脑组织缺血早期NMDAR即被激活,大量Ca2+内流致细胞内钙超载,进一步引起自由基的产生,乌司他丁降低了此种损伤,但确切的机制还有待进一步的研究。
乌司他丁减轻脑缺血再灌注损伤的机制可能与抑制NMDAR1表达有关,但不同剂量、不同时间点对神经细胞的作用以及远期效果还待进一步的研究,从而为其临床应用提供科学依据。
