牛乳酪蛋白基因多态性研究进展
2021-02-25赵烜影刘振民雍靖怡穆海菠
赵烜影,刘振民,雍靖怡,穆海菠,李 楠*
(乳业生物技术国家重点实验室,上海乳业生物工程技术研究中心,光明乳业股份有限公司乳业研究院,上海 200436)
牛乳是犊牛最重要的食物,也是人类补充蛋白质和微量元素的常见来源。牛乳中有2大类蛋白质:乳清蛋白和酪蛋白(casein,CN),乳清蛋白由β-乳球蛋白(β-lactoglobulin,β-Lg)、α-乳白蛋白(α-lactalbumin,α-La)、免疫球蛋白、糖肽、牛血清白蛋白和次要蛋白,如乳过氧化物酶、溶菌酶和乳铁蛋白组成[1];CN约占牛乳蛋白的76%~86%,是由乳腺上皮细胞合成的一种含有大量钙和磷的蛋白,以胶束形式存在于牛乳中。酪蛋白含有人体所必需的8 种氨基酸。牛乳含有4 种酪蛋白:αs1-CN(CSN1S1,占总酪蛋白的39%~46%)、αs2-CN(CSN1S2,占总酪蛋白的8%~11%)、β-CN(CSN2,占总酪蛋白的25%~35%)和κ-CN(CSN3,占总酪蛋白的8%~15%)[2]。这4 种酪蛋白通过磷酸化、糖基化和水解会产生一部分新的酪蛋白,如αs-CN通过磷酸化、κ-CN通过糖基化以及β-CN经过水解,会生成γ-酪蛋白和多肽等[3]。酪蛋白多态性被认为会影响乳产量、乳的理化特性和营养成分、乳制品加工特性以及人类的营养而得到学者们的重视。
1 酪蛋白基因多态性
超过95%反刍动物乳中的酪蛋白都是通过6 个结构基因编码形成的,在6号染色体上4 个酪蛋白基因组成了一个250 kb的片段,如图1所示。而水牛乳的4 种酪蛋白则同样由4 个在水牛7号染色体上呈簇状分布的酪蛋白基因编码[5]。Aschaffernburg等[6]以β-Lg为对象,找出了2 种基因型,发现了乳蛋白的基因多态性。越来越多的学者开始研究乳蛋白基因多态性,乳蛋白基因多态性在不同牛种之间存在明显差异,为了适应当地饲育条件而杂交培育出的本地特色奶牛则进一步丰富了牛乳酪蛋白的基因多态性。
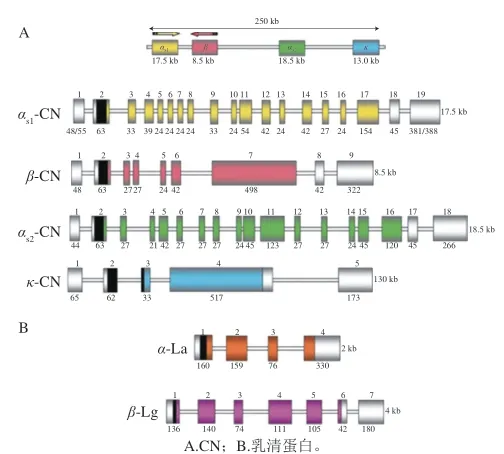
图 1 主要乳蛋白编码转录单位的结构组织[4]Fig. 1 Structural organization of major milk protein coding transcription units[4]
在众多研究中发现会出现同一种等位基因在不同物种中命名不同的情况,这可能与检测技术不同相关。尽管在DNA水平上的鉴定可以发现很多单核苷酸多态性(single nucleotide polymorphism,SNP)位点,但是这些突变位点可能是没有更改氨基酸序列的同义突变位点,进而在蛋白质水平上无法检测出新的蛋白质亚型。Farrell等[1]提出的对普通牛、瘤牛、牦牛、爪哇野牛乳蛋白基因型命名法更加适用于常见的乳蛋白等位基因。目前已被报道的牛乳蛋白基因有A、B、C、D、E、F、G、H、I、J 10 种CSN1S1等位基因;A、B、C、D、E 5 种CSN1S2等位基因;A1、A2、A3、B、C、D、E、F、G、H1、H2、I、J、K、L 15 种CSN2等位基因和A、B、B2、C、D、E、F1、F2、G1、G2、H、I、J 13 种CSN3等位基因[7-9]。
1.1 α-CN基因多态性
αS1-CN由214 个氨基酸组成的肽链和1 个含有15 个氨基酸的前导链组成[10]。编码αS1-CN的CSN1S1基因全长17 508 bp,有18 个内含子及19 个外显子[11]。αS1-CN在磷酸钙的运输、酪蛋白胶束的形成以及酪蛋白从内质网传递到高尔基体中起到关键作用。CSN1S1基因多态性由于与牛乳的营养价值和加工特性相关而得到广泛关注[12]。CSN1S1的B等位基因最为常见;在DNA序列上的26 139 bp处由C突变为T的A等位基因在埃及、意大利等地水牛中被多次报导;C等位基因在利木赞牛、瑞士褐牛、弗莱维赫牛等杂交品种中发生频率最高(>10%)[2]。Fan Xinyang等[11]在沼泽水牛中发现了由c.609T>G产生的D等位基因。I等位基因被证实是由c.296A>T产生[1]。2013年,在吉尔牛和短角牛中发现了c.543G>T,并将其命名为J等位基因[9],至此CSN1S1基因增加为10 种。除B、C等位基因外,其余等位基因仅在少数品种中出现。
编码αS2-CN的CSN1S2基因全长18 480 bp,有18 个外显子和17 个内含子[13]。CSN1S2基因多态性被认为与泌乳性能有关。目前已报道的CSN1S2等位基因共有A、B、C、D、E 5 种,大都集中在第3、第6及第8个外显子中。等位基因B、C、D、E均被认为是由A等位基因突变而来,A等位基因外显子3的第17位碱基发生了C→T的替换,产生了B等位基因;外显子6的第2位碱基发生了A→G的替换,产生了C等位基因;外显子8的第26位碱基发生了G→T的替换,产生了D等位基因;外显子3的第13位碱基发生了C→T的替换,产生了E等位基因[13]。
1.2 β-CN基因多态性
β-CN是一种疏水性球状蛋白,由209 个氨基酸残基组成,分子质量为25.382 kDa,牛乳中的β-CN以完全磷酸化形式存在[14]。编码β-CN的CSN2基因全长8.5 kb,由9 个外显子和8 个内含子组成[15]。β-CN的基因多态性被认为与Ⅰ型糖尿病、心血管疾病、自闭症、精神分裂和免疫缺陷等多种疾病相关[16]。目前已被报道的CSN2等位基因共有15 种(A1、A2、A3、B、C、D、E、F、G、H1、H2、I、J、K、L)[17]。CSN2等位基因及其发生频率因检测品种、样本量及地域的差异而存在较大差异。意大利北部荷斯坦牛的检测结果显示,A1发生频率为0.371,A2发生频率为0.546,而美国对于荷斯坦牛的检测结果与上述结果相反,A1的发生频率(0.31~0.66)超过了A2(0.24~0.62)[16-17]。但总的来说,在现有研究结果中,A1和A2等位基因以超过0.4的发生频率最为常见[18]。B等位基因的发生频率为0.01~0.10[17]。I等位基因发生频率在大部分品种中均低于0.03,但在意大利红白花牛(0.14)和德国荷斯坦-弗里森杂交牛(0.192)中显示出较高的频率[19]。有学者在韩国本地牛类中检测到A4等位基因,但尚未发现其SNP位点,D等位基因只在瘤牛中被发现,E等位基因只在皮埃蒙特品种中被发现[4]。A3、C、F、G等位基因较为罕见,发生频率均低于0.01[20]。目前尚未有关于H1和H2等位基因发生频率数据。
1.3 κ-CN基因多态性
κ-CN多存在于酪蛋白胶束的表面,对维持胶束的稳定性起到重要作用[21]。κ-CN含有半胱氨酸和蛋氨酸2 种含硫氨基酸,是一种糖基化酪蛋白[1],由编码190 个氨基酸的肽链组成,其中21 个氨基酸形成信号肽前导链,编码κ-CN的CSN3基因全长573 pb,含有5 个外显子和4 个内含子,大多数编码序列包含在第4个外显子中[22]。CSN3基因首次被证实是在1964年,迄今为止已发现13 个等位基因(A、B、B2、C、D、E、F1、F2、G1、G2、H、I和J)及1 个同义变体(A1)[23]。目前已知的等位基因都应该是A等位基因由于氨基酸的缺失或置换等突变引起的,其中A、B等位基因因与牛乳产量及凝乳特性有关而被研究人员重视[11]。在荷斯坦牛中A等位基因的发生频率为0.736,远高于B等位基因的0.186,但是在娟姗牛中B等位基因的发生频率(0.692)超过了A等位基因(0.308)[9]。A等位基因具有136Thr(ACC)/148Asp(GAT),而B等位基因具有136Ile(ATC)/148Ala(GCT),E等位基因将氨基酸155Ser(AGC)更改为155Gly(GGC)[4]。G等位基因比A、B等位基因少见,仅在德国黑白花牛、荷斯坦-弗里斯兰牛、捷克弗莱维赫牛、艾尔郡牛和瑞典红牛等少数品种中发现过[24]。
2 基因多态性检测方法
乳蛋白的基因多态性主要是由氨基酸的缺失或置换、糖基化、磷酸化位点的差异引起的,目前,关于基因多态性的检测大都是在蛋白水平和DNA水平上进行。蛋白水平检测常见的方法包括毛细管区带电泳、反相高效液相色谱(reversed-phase high-performance liquid chromatography,RP-HPLC)、等电聚焦电泳或尿素-聚丙烯酰胺凝胶电泳[25-28]。最近,液相色谱-串联质谱或液相色谱-高分辨率质谱等更强大的技术已被成功地用于蛋白质变体的鉴定和定量[29-30],主要包括等位基因特异性聚合酶链式反应(allele specific polymerase chain reaction,AS-PCR)、限制性片段长度多态性PCR(restriction fragment length polymorphism PCR,RFLP-PCR)、单链构象多态性PCR(single strand conformation polymorphism PCR,SSCP-PCR)、创造酶切位点PCR(amplification created restriction site PCR,ACRS-PCR)等方法,近年来还出现了TaqMan法及焦磷酸测序等技术[31-32]。
2.1 蛋白质水平检测方法
HPLC法虽然不是最常见的蛋白质组学分析技术,但适用于初始阶段分离多种蛋白质混合物。RP-HPLC法通常采用十八烷基-硅胶固定相和极性强于固定相的低离子强度酸性有机流动相。随着流动相有机溶剂浓度的改变,蛋白质亲硅醇基效应和疏水性会影响到蛋白质的保留时间。不同亚型的酪蛋白由于-NH2和-COOH等极性和氢键作用较强的基团含量及位置的不同,保留时间不同,而被分离。C8色谱柱属于短链且不具有支链的键合固定相,其保留性弱于C18,因而更适用于出峰时间差异较小的酪蛋白亚型之间的测定。刘亚楠等[33]通过RP-HPLC法成功检测到中国水牛4 种酪蛋白的不同亚型,发现κ-CN出现了3 种不同的峰,即A、B、C 3 种等位基因,αS1-CN、αS2-CN及β-CN各出现了2 个不同的峰。陈晨等[34]利用RP-HPLC法在杂交水牛的κ-CN中发现了新的D等位基因。操作简便、耗时短、回收率高、可以检测出沉默变异体等优点使HPLC法成为检测酪蛋白多态性的有效方法。
液相色谱-质谱联用方法是将液相色谱分离后的目标蛋白质通过电离的方式引入质谱,得到蛋白质不同亚型特异性肽段的质谱图。将测定出的单同位素质量与已知的蛋白质亚型单同位素质量进行对比,来鉴定蛋白质的多态性。Nguyen等[31]通过使用与Q-Exactive质谱仪偶联的Ultimate 3000 UPLC成功检测到越南黑白花荷斯坦F2(HF2)乳样中β-CN的3 个亚型,在85 头HF2乳样中检测到3 个单同位素质量分别为24 008.143 0、23 968.167 0、23 950.226 0 Da的亚型,分别与β-CN亚型中A1、A2和I的理论单同位素质量24 008.161 6、23 968.151 6、23 950.191 6 Da匹配。
不同的电泳技术则是通过蛋白质亚型在电泳或等电聚焦过程中迁移率的差异进行检测,可以对多种乳蛋白进行同时检测。Caroli等[27]开发了一种将等电聚焦电泳后胶片浸泡在三氯乙酸中,将β-CN亚型固定化的定量方法,避免了考马斯亮蓝染色过程β-CN与β-Lg重叠,该方法不仅成功检测出β-CN的A1和A22 种亚型,还能够对样品中的β-CN亚型进行定量分析。冯慿等[35]建立的毛细管区带电泳方法可以对液态乳和乳粉中β-CN亚型进行定性及定量分析,成功检测出A1及A22 种亚型。Duarte-Vázquez等[28]通过尿素-聚丙烯酰胺电泳成功检测出荷斯坦及娟姗牛β-CN的A1、A22 种亚型。
蛋白质水平上的检测方法由于能够更加直观验证酪蛋白的基因型而在实际中应用更加广泛。
2.2 DNA水平检测方法
焦磷酸测序作为一种新型的酶联级联测序技术适用于对已知短序列的测序分析,其通过引物与模板DNA退火后,在DNA聚合酶、三磷酸腺苷(adenosine triphosphate,ATP)硫酸化酶、荧光素酶和ATP双磷酸酶4 种酶的协同作用下,将引物上dNTP的聚合与荧光信号的释放偶联,检测荧光的释放和强度,达到实时测定DNA序列的目的[36]。可以大通量、低成本、实时、快速、直观地进行SNP测定。任大喜等[37]采用焦磷酸测序法分析中国荷斯坦牛、娟姗牛和水牛的乳蛋白基因多态性,并找出了水牛κ-CN的特异性多态位点。
AS-PCR方法的原理是将突变碱基设计在引物3’末端,在PCR中若突变碱基与模板互补,获得特异扩增条带,则表明被测基因含有该种突变[38]。Gholami等[39]使用AS-PCR技术扩增了1 个854 bp的片段,成功鉴定出伊朗荷斯坦牛、娟姗牛、当地杂交奶牛β-CN的A1、A2等位基因,并测定出A1等位基因的发生频率为荷斯坦牛0.5、娟姗牛0.51、西斯塔尼牛0.54、塔莱西牛0.49、马赞达拉尼牛0.46。Firouzamandi等[40]同样利用AS-PCR技术在荷斯坦Sarabi牛中检测到β-CN的A1、A2等位基因。
RFLP-PCR技术是将PCR扩增后的DNA用特异性内切酶切分成长短不一的片段,并在凝胶电泳上分辨。因为不同等位基因的限制性酶切位点分布不同,会产生不同长度的DNA片段条带。Cinar等[41]利用该方法设计正向引物5’-ATCATTTATGGCCATTCCACCAAAG-3’、反向引物5’-GGCCATTTCGCCTTCTCTGTAACAGA-3’,成功测定了安那托利亚水牛κ-CN的B等位基因。Jawane等[42]使用CASB67R和CASB122L引物扩增印度瘤牛β-CN基因外显子VII的一部分,并用TaqI限制酶对251 bp的PCR扩增序列进行限制性酶切,得到3 种基因型:A1A1(213、38 bp)、A1A2(251、213、38 bp)和A2A2(251 bp)。
SSCP-PCR技术的基本原理是将PCR扩增后的DNA片段经变性形成单链DNA,单个碱基的差异会在中性聚丙烯酰胺凝胶电泳时形成不同的立体构象,改变泳动速率,产生不同的条带。Chiatti等[43]运用此方法成功鉴定出山羊乳αS2-CN的A、B、C、E共4 个等位基因,并通过等电聚焦在蛋白质水平上验证,他们认为SSCP-PCR技术相比RFLP-PCR技术在DNA水平上检测CSN1S2等位基因会更加快捷、平价。Hamza等[44]利用SSCP-PCR技术设计正向引物5’-CTAAATCTGGCATAAAAGTA-3’、反向引物5’-AATCACGGACTAAATAA-3’的引物对,成功检测出中国荷斯坦奶牛κ-CN的C、T等位基因。
TaqMan荧光探针是一种寡核苷酸探针,其5’末端携带荧光基团,如FAM、TET、VIC、HEX等,3’端携带猝灭基团,如TAMRA、BHQ等。其原理为:用1 对双标记的TaqMan荧光探针分别针对双等位SNP的不同等位基因,只有完全匹配的探针可以扩增出其对应的等位基因。若只有VIC荧光信号明显增强表明为等位基因1,若只有FAM荧光信号明显增强表明为等位基因2,当VIC与FAM荧光信号均明显增强则为等位基因1与2的杂合型。而新型TaqMan-MGB(minor groove binder)探针使该技术既可进行基因定量分析,又可分析基因突变(SNP)[45]。Kusza等[46]利用该技术发现罗马尼亚本土扎克尔绵羊CSN2基因的A等位基因发生频率为0.009 8。
3 酪蛋白基因多态性的应用
3.1 酪蛋白基因多态性在生产加工中的应用
作为奶牛的重要经济指标,泌乳性能由多个基因控制,乳蛋白基因则是影响泌乳性能的主要基因,乳蛋白中编码酪蛋白和β-Lg的基因对奶牛的泌乳性能影响最大。梁艳等[47]对智利进口的荷斯坦牛酪蛋白多态性进行研究发现,β-CN多态性对日产奶量有极显著影响,AB亚型个体的日产奶量极显著高于AA亚型(P<0.01)。杨雪瑶等[48]研究宁夏地区荷斯坦奶牛发现,含κ-CN的AA基因型奶牛的产奶量显著高于CA基因型(P<0.05),极显著高于CC基因型(P<0.01),说明κ-CN的A等位基因有利于提高日产奶量。Morkuniene等[49]对立陶宛牛群κ-CN多态性进行研究同样发现,A等位基因有利于提高日产奶量。
除了影响奶牛泌乳性能以外,酪蛋白多态性对牛乳营养成分的影响更为显著。Gurses等[23]研究κ-CN发现,含AB亚型的荷斯坦牛乳蛋白质及乳糖含量显著高于AA及AE亚型;含A等位基因的娟姗牛乳脂含量显著低于含B等位基因。也有研究显示,水牛β-CN的B等位基因与乳蛋白含量成正比,而A等位基因则与乳脂含量成正比[50]。黄改奶牛αs2-CN的A等位基因与乳脂及乳蛋白含量成正相关[13]。Li Shanshan等[51]研究中国水牛的酪蛋白遗传多态性与乳蛋白组成之间的关联,发现κ-αs1-β复合基因型中,BB-AB-AA型和AB-AB-AA型分别对κ-CN和αs2-CN含量有显著影响。Day等[52]研究酪蛋白多态性对乳蛋白胶束大小的影响,发现κ-CN的AA亚型酪蛋白胶束显著大于AB及BB亚型,并且小胶束酪蛋白((151.0±2.2) nm)的糖基化比例更高,即含B等位基因的酪蛋白更容易糖基化;同时还发现小胶束酪蛋白中β-CN出现基因突变的频率更高。陈晨等[34]研究中国水牛的酪蛋白多态性发现,水牛αs1-β-κ-CN复合基因为ABBB-AB型,κ-CN的C等位基因均与乳蛋白粒度存在显著相关性,αs1-CN和κ-CN基因型为A等位基因的蛋白粒度大于基因型为其他等位基因的蛋白。
酪蛋白多态性对牛乳加工性能的影响也是学者们的研究重点。黄丽[53]、李玲[54]等研究水牛αs1-CN对马苏里拉干酪及类蒙特利杰克半硬质干酪加工及品质的影响,发现水牛αs1-CN多态性对马苏里拉干酪组成影响显著,αs1-CN为AB亚型的水牛乳制作的马苏里拉干酪在脂肪、蛋白质含量以及干酪硬度、弹性、胶黏性、咀嚼性等指标方面都优于BB亚型;但是αs1-CN为BB亚型的水牛乳制作的类蒙特利杰克半硬质干酪在各项指标上均显著优于AB亚型。Zicarelli等[55]研究发现,含有意大利地中海水牛αs1-CN的B等位基因以及κ-CN的F1等位基因的牛乳凝乳效率显著高于含其他等位基因的牛乳。Nguyen等[56]研究奥克兰当地牧场奶牛β-CN的A1与A2等位基因对牛乳中钙分布、酸凝固、起泡特性及酸乳凝胶微观结构的影响,发现含有β-CN A2等位基因的牛乳中游离Ca2+含量显著高于含A1等位基因的牛乳,泡沫形成能力增强,需要更长的时间才能凝固;A2A2亚型酸乳的凝胶强度较弱,微观结构较多孔,表明该凝胶更细腻,在外部机械力的作用下容易破裂和变形,因此,在处理和运输A2A2亚型酸乳产品期间可能需要格外小心,然而从健康和营养的角度来看,A2A2亚型酸乳较软,而较弱和较多孔的凝胶有助于蛋白质更快地分解,在胃中酸性条件下更好被消化。在对中国水牛β-CN基因多态性的研究过程中发现,体外消化实验表明,AB亚型消化最快,其次为AA亚型,BB亚型最难消化[50]。
3.2 酪蛋白基因多态性对人体健康的潜在影响
关于酪蛋白多态性对人体健康的影响,学者们大都聚焦在β-CN的A1与A2等位基因,这可能与β-CN含量及A1和A2等位基因发生频率高有关。β-CN的A1等位基因在人体内经过胃蛋白酶的消化会产生一种阿片肽——β-酪啡肽7(β-casomorphin 7,BCM7),其产生机制见图2。BCM7被认为与Ⅰ型糖尿病、缺血型心脏病、精神分裂、自闭症、婴儿猝死综合征、动脉粥样硬化、冠状动脉性心脏病、孤独症谱系障碍等疾病有关[16,32,57]。
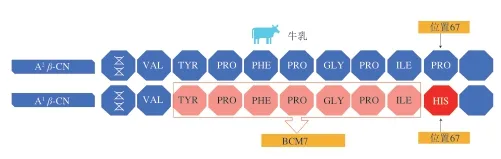
图 2 BCM7产生机制[57]Fig. 2 BCM7 generation mechanism[57]
多位学者研究发现A1等位基因的摄入量与Ⅰ型糖尿病的发病率有显著相关性;对BioBreeding(BB)大鼠的动物实验也显示,A1等位基因有诱发Ⅰ型糖尿病的作用[58]。关于诱发机制目前存在2 种假说:1)含A1等位基因的β-CN会刺激T免疫细胞产生免疫反应及抗体免疫反应,这会促进Ⅰ型糖尿病的发病;2)类似于BCM-7的阿片类药物可能会干扰代谢过程,包括血糖水平的调节和胰岛素的产生,进而导致Ⅰ型糖尿病,动物实验也证实了阿片肽类药物的共同给药受体拮抗剂纳洛酮有效减弱了A1等位基因的作用[58]。
对食物摄入的不良反应具有非常多样的病因和症状,在牛乳中主要表现为乳蛋白过敏和乳糖不耐症。乳蛋白过敏是最常见的食物过敏症状,对儿童的影响较大,尤其是新生儿。一般认为BCM7会刺激T免疫细胞,进而引起乳蛋白过敏,β-CN的A2等位基因则会降低致敏乳蛋白的表达[59]。当然,除了β-CN以外,β-Lg与α-La也是乳蛋白过敏的重要过敏原。乳糖不耐症则是因为体内缺乏乳糖酶,乳糖在肠道中分解为乳酸,破坏了肠道酸碱平衡,进而引起腹泻,是一种多发于亚洲地区的先天性遗传疾病,部分人体内的乳糖酶活力会随着年龄的增加而降低。He Mei等[60]通过临床实验研究发现,含A1β-CN牛乳会降低乳糖酶活性,并增加胃肠道症状,而含A2β-CN牛乳可减轻急性乳糜泻引发的胃肠道不适症状。目前尚未有直接的证据解释β-CN的A1等位基因对肠胃影响的机理。
氧化代谢是维持细胞生命活动的重要功能之一,新陈代谢失调会产生许多活性氧和自由基,引起氧化应激。过氧化物自由基、羟自由基、超氧自由基和过氧化氢等自由基不仅会引起蛋白变质,还会损害生物系统。由于蛋白质和多肽具有特定的或非特异性的生物学机制,如使活性氧失活、清除促氧化的金属离子、降低氢过氧化物水平以及改变系统的内部特性,因此蛋白质和多肽具有抗氧化潜力;对中国水牛酪蛋白多态性与其抗氧化性的研究发现,AA亚型β-CN的1,1-二苯基-2-三硝基苯肼自由基清除能力优于BB亚型,BB亚型的羟自由基清除能力和还原能力优于AA亚型[50]。
4 结 语
酪蛋白作为牛乳中的重要组成部分,具有极高的营养价值。4 种酪蛋白均存在基因多态性,这些多态性对牛乳的组成成分、牛乳品质、牛乳的生产加工特性以及乳制品的营养、口感、品质具有重要影响。加强对酪蛋白多态性的基础研究可以为保护牛的基因多态性、奶牛的定向育种以及精准设计功能性乳制品提供理论基础,引导中国乳业向高附加值、高收益多样化发展。