BMP4蛋白及其受体在不同发育期大鼠胃肠道的表达分布
2021-02-10孟宪博韩士举梁世洋王景杰
孟宪博,林 强,韩士举,梁世洋,王景杰
(空军军医大学唐都医院消化内科,西安 710038;*通讯作者,E-mail:jingjie@fmmu.edu.cn)
骨形态发生蛋白(bone morphogenetic protein, BMP)是转化生长因子β(transforming growth factor-β)蛋白超家族成员,与骨和软骨形态发生、修复相关[1]。随着研究的进展发现,BMP除参与骨形态发生调节外,还参与诸多骨外组织的发育分化及功能维持[2],其中BMP4在消化道发育、组织分化和功能维持中起到重要作用[3,4]。但目前关于BMP4及其受体在不同发育阶段大鼠消化道分布表达尚不清楚,对于其在不同消化道器官功能维持和分化发育的作用机制仍不明确。
前期研究表明,在胚胎消化道发育过程中,BMP4协同Hedghog通路相关蛋白参与消化道环形肌和纵行肌分化发育,且对于正常胚胎发育不可或缺[3]。除参与胚胎消化器官发育外,出生后正常消化器官稳态的维持同样有BMP信号的参与调节,在炎症、肿瘤和其他消化道疾病的影响下可能会产生BMP表达和分布异常[5-7]。有临床研究表明,在许多消化道疾病和消化道肿瘤中,BMP4呈现出异常的表达水平和分布特点[2]。有研究报道BMP4与食管腺癌的发生密切相关,且BMP4基因突变与结直肠癌的发生同样具有很高的相关性[8]。
因此,我们推测不同发育阶段大鼠消化道器官中BMP4及其受体可能有分布和表达的差异性,或与正常消化道黏膜腺体及平滑肌功能维持密切相关。
为此本研究拟通过形态学和分子生物学方法,研究BMP4及其受体BMPR1B在不同发育阶段大鼠消化道中的表达分布情况,特别关注BMP4在不同发育阶段的表达差异,进而探讨BMP4在胃肠发育调节以及胃肠疾病中的意义。
1 材料与方法
1.1 实验动物
5日龄、3周龄及8周龄雄性SD大鼠各6只,SPF级。5日龄大鼠体质量10-12 g,3周龄大鼠体质量50-60 g,8周龄大鼠体质量250-300 g,由北京维通利华实验动物技术有限公司提供,实验动物生产许可证号:SCXK(京)2016-0011。
1.2 主要试剂、仪器
兔抗BMP4抗体(美国Abcam公司),兔抗BMPR1B抗体(美国Abcam公司),Alexa Fluor 594标记驴抗大鼠IgG抗体(美国Jackson公司),Alexa Fluor 488标记驴抗兔IgG抗体(美国Jackson公司),HRP酶标记的羊抗兔IgG抗体(美国Santa Cruz公司),HRP酶标记的羊抗小鼠IgG抗体(美国Santa Cruz公司),小鼠抗GAPDH抗体(中国Servicebio);Nikon eclipse Ci荧光显微镜(日本NIKON公司);冷冻切片机(德国Leica公司);电泳仪和电泳槽(美国Bio-Rad公司)。
1.3 胃肠道组织标本制备
用异氟烷诱导麻醉大鼠并维持,打开胸腔用预冷0.01 mol/L PBS溶液灌流心脏,打开腹腔观察肝叶褪色后取胃、十二指肠及结肠用0.01 mol/L预冷PBS反复冲洗。分别将用于Western blot冲洗干净后的胃、十二指肠和结肠组织剪碎,加入含蛋白酶抑制剂及磷酸酶抑制剂的RIPA裂解液,匀浆器匀浆,冰上裂解30 min,4 ℃ 12 000 r/min离心15 min,吸取上清,用BCA法蛋白定量。用于免疫荧光的8周龄大鼠消化道组织4%多聚甲醛固定各节段消化道组织6-8 h,将固定好的组织置于25%蔗糖溶液,4 ℃浸泡过夜脱水沉底,包埋后冰冻切片机上连续切片,切片厚10 μm。
1.4 间接免疫荧光法检测大鼠胃肠道BMP4及BMPR1B分布
将制备好的8周龄大鼠胃体、结肠升段组织冰冻切片,用含1% BSA+0.5%Triton的PBS液封闭40 min,加一抗(BMP4抗体按1 ∶100稀释,α-SMA抗体按1 ∶200稀释)。一抗4 ℃孵育过夜。经PBST漂洗5 min×3次,后加二抗(Alexa Fluor 488荧光标记二抗或,Alexa Fluor 594荧光标记二抗均按1 ∶1 000稀释)室温孵育2 h,DAPI工作液室温孵育5 min衬核。PBST漂洗后,50%缓冲甘油封片,荧光显微镜下观察染色后的胃体及结肠升段组织切片。
1.5 Western blot检测大鼠胃肠道BMP4及BMPR1B相对表达水平
对5日龄、3周龄、及8周龄的大鼠用Western blot法半定量检测胃、十二指肠、结肠组织BMP4及BMPR1B蛋白表达水平,取制备好的大鼠胃体、十二指肠及升结肠蛋白,绝对定量后用含蛋白酶及磷酸酶抑制剂的RIPA调整蛋白浓度至一致。分别加入loading buffer,金属浴95 ℃ 10 min。对制备好的蛋白样品进行SDS-PAGE凝胶电泳、常规转膜。将转好的PVDF膜,用TBST配制的5%脱脂牛奶室温封闭1 h,封闭后TBST洗膜10 min×3次,再加兔抗BMP4/BMPR1B抗体(按1 ∶1 000稀释)4 ℃孵育过夜。TBST漂洗15 min×2次。加HRP酶标记的羊抗兔IgG二抗,室温孵育1 h。TBST漂洗后,进行ECL化学发光反应。
1.6 统计学分析
Western blot条带灰度值用Image pro plus软件测量,并用SPSS21.0统计软件进行单因素方差分析,Bonferroni法进行多个样本均数间多重比较,P<0.05为差异有统计学意义。
2 结果
2.1 BMP4阳性细胞在胃肠道的组织分布特点
采用间接免疫荧光方法双标α-SMA蛋白与BMP4蛋白,结果显示:BMP4广泛分布于8周龄大鼠胃及结肠全层。
胃腺体、黏膜肌层及肌层均可见大量BMP4分布,而黏膜下层未见明显BMP4分布。相较于胃腺体,黏膜肌层及肌层BMP4分布更加广泛,且在胃体外层纵行肌与中层环形肌交界处BMP4分布相对更加聚集(见图1)。
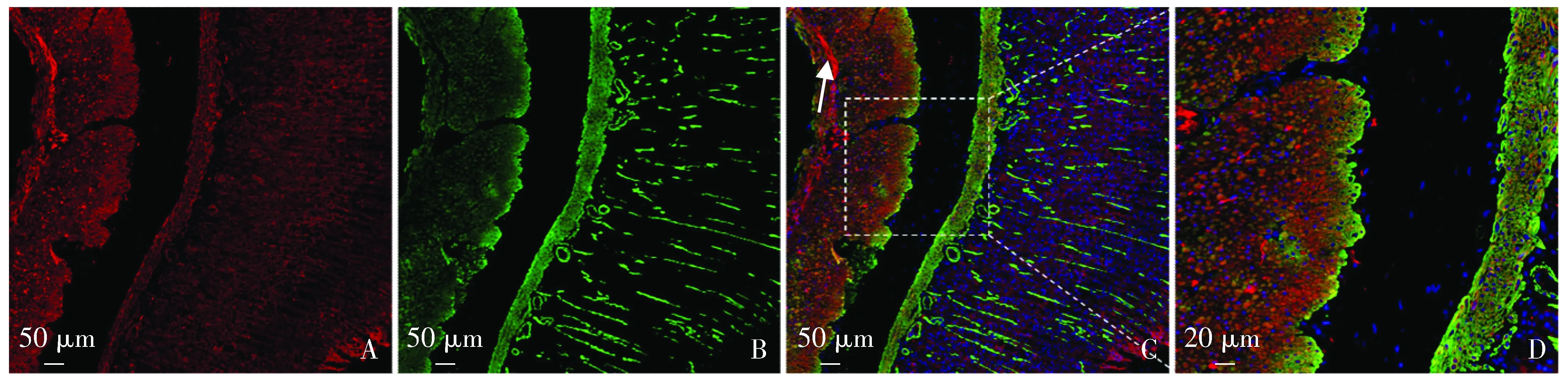
A.胃体BMP4分布(红);B.胃体平滑肌细胞(α-SMA标记,绿);C,D.BMP4和α-SMA双标图1 8周龄大鼠胃体BMP4分布特点Figure 1 Distribution of BMP4 in stomach of 8-week old rats
在结肠中,BMP4蛋白多在分泌后聚集于平滑肌层,黏膜层可见腺体细胞中由少量BMP4蛋白聚集,黏膜下层未见明显染色。在结肠升段平滑肌层可见环形肌及纵行肌中广泛分布BMP4,且在环形肌和纵行肌交界处,BMP4分布更加密集(见图2A-D)。
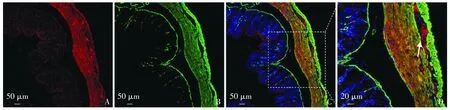
A.结肠升段BMP4分布(红);B.结肠平滑肌细胞(α-SMA标记,绿);C,D.结肠BMP4和α-SMA双标图2 8周龄大鼠结肠升段BMP4分布特点Figure 2 Distribution of BMP4 in ascending colon of 8-week old rats
2.2 BMPR1B阳性细胞在胃肠道的组织分布特点
结果显示:BMP4蛋白受体BMPR1B在8周龄胃体和结肠升段中均有分布,且在胃体和结肠升段分布呈现一定的特异性。
在胃体中,共标平滑肌α-SMA蛋白与BMPR1B可见,BMPR1B在胃体黏膜层腺体细胞中广泛分布,黏膜下层未见明显分布,肌层中散在分布BMPR1B。在胃体肌层中,外侧纵行肌与中层环形肌交界处可见较多点状分布BMPR1B(见图3)。
在大鼠结肠升段中,双标α-SMA蛋白与BMPR1B可见,黏膜层和结肠肌层中有BMPR1B分布,黏膜下层未见明显染色。黏膜层中,BMPR1B多分布在黏膜腺体细胞,黏膜肌层未见明显分布。结肠升段平滑肌层中可见平滑肌间散在分布BMPR1B(见图4)。
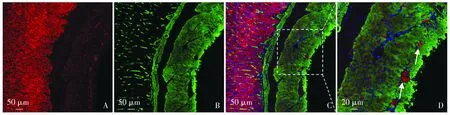
A.胃体BMPR1B阳性细胞(红);B.胃体平滑肌细胞(α-SMA标记,绿);C,D.胃体BMPR1B和α-SMA双标图3 大鼠胃体BMPR1B分布特点Figure 3 Distribution of BMPR1B in stomach of rats
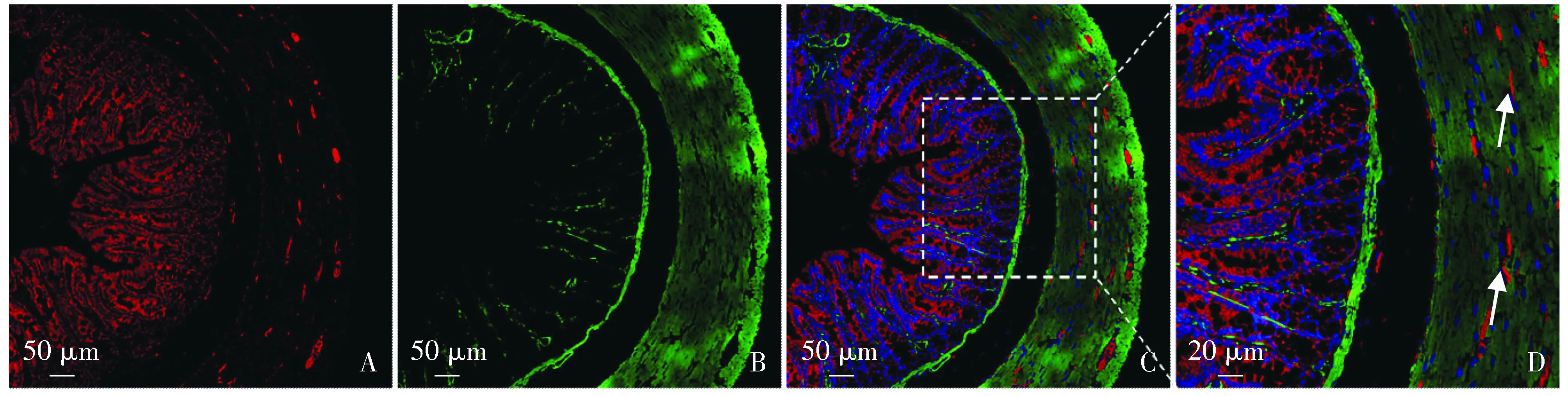
A.结肠BMPR1B阳性细胞(红);B.结肠平滑肌细胞(α-SMA标记,绿);C,D.BMPR1B和α-SMA双标图4 大鼠结肠升段BMPR1B分布特点Figure 4 Distribution of BMPR1b in rat ascending colon
2.3 BMP4在胃肠道的时空表达特点
结果示BMP4在胃、十二指肠、结肠均有表达。
5日龄、3周龄与8周龄大鼠胃体中BMP4蛋白表达水平差异有统计学意义(F=23.6,P<0.05),且5日龄、3周龄大鼠胃体中BMP4表达水平分别高于8周龄(P<0.05)。5日龄和3周龄大鼠十二指肠BMP4表达水平分别高于8周龄大鼠,差异有统计学意义(P<0.05),而5日龄与3周龄大鼠结肠中BMP4表达水平未见明显差异(P>0.05),但表达水平与8周龄大鼠相比差异有统计学意义(F=58.0,P<0.05)。
对于同一发育阶段大鼠,胃、十二指肠与结肠BMP4表达水平差异有统计学意义(F5 d=40.2,P<0.05;F3周=93.8,P<0.05;F8周=13.3,P<0.05)。在5日龄和3周龄大鼠中,胃体与十二指肠BMP4表达水平均高于结肠,差异有统计学意义(P<0.05,见图5)。
2.4 BMPR1B在胃肠道的时空表达特点
结果示BMPR1B在胃、十二指肠、结肠均有表达(见图6)。
对于不同发育阶段大鼠,3周龄大鼠胃体BMPR1B表达水平高于5日龄及8周龄大鼠,差异有统计学意义(P<0.05),且3周龄大鼠十二指肠BMPR1B表达水平同样高于5日龄及8周龄大鼠,差异有统计学意义(P<0.05)。5日龄、3周龄与8周龄大鼠结肠BMPR1B表达水平差异有统计学意义(F=45.1,P<0.05)。
对于同一发育阶段大鼠不同消化道部位,3周龄与8周龄大鼠胃体BMPR1B表达水平均高于结肠,差异有统计学意义(P<0.05)。5日龄大鼠胃体、十二指肠与结肠BMPR1B表达水平差异有统计学意义(F=38.4,P<0.05)。
3 讨论
骨形态发生蛋白属于转化生长因子β家族,包含20种结构类似的蛋白[9]。在细胞增殖、分化和凋亡过程中,骨形态发生蛋白家族起到至关重要的作用,如敲除BMP2/4基因对于小鼠胚胎来说具有致死性[4,10]。BMP通过与其受体结合,形成Ⅰ/Ⅱ型丝苏氨酸激酶受体。Ⅰ型和Ⅱ型受体结构类似,不同的BMP蛋白较为特异地选择结合不同类型的BMP受体。激活受体后,BMP通过经典通路或非经典通路磷酸化下游Smad1/5/8蛋白,参与细胞内信号转导[11],同时与其他信号通路间同样存在交流[12,13]。
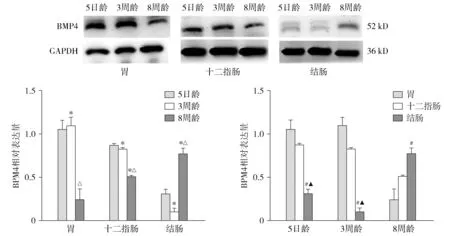
与5日龄比较,*P<0.05;与3周龄比较,△P<0.05;与胃体比较,#P<0.05;与十二指肠比较,▲P<0.05图5 BMP4蛋白在不同发育阶段大鼠胃、十二指肠、结肠的表达水平差异Figure 5 Expression of BMP4 protein in stomach, duodenum and colon of rats with different developmental stages
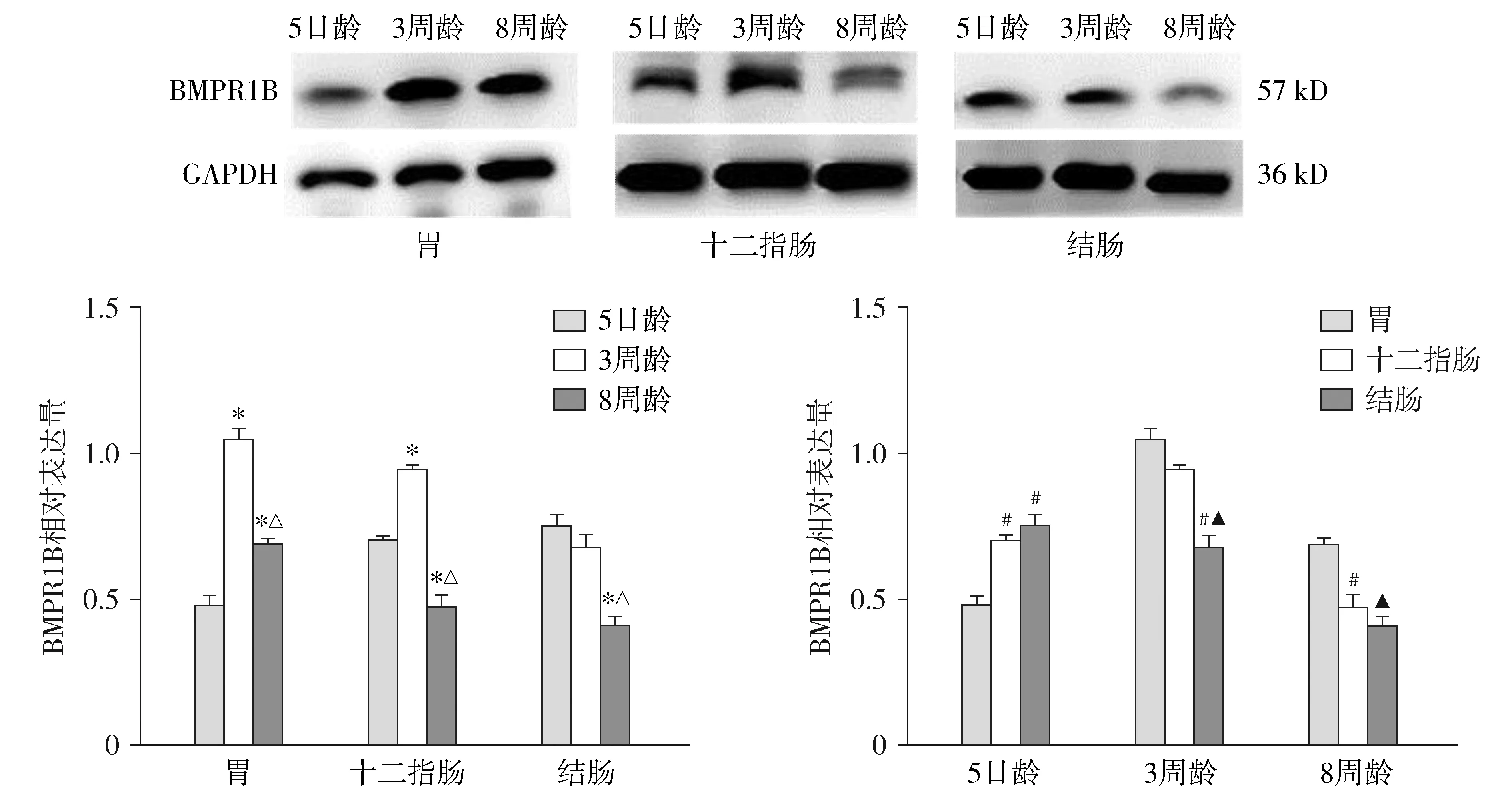
与5日龄比较,*P<0.05;与3周龄比较,△P<0.05;与胃体比较,#P<0.05;与十二指肠比较,▲P<0.05图6 BMPR1B蛋白在不同发育阶段大鼠胃、十二指肠、结肠的表达水平差异Figure 6 Expression of BMPR1B protein in stomach, duodenum and colon of rats with different developmental stages
临床研究提示,骨形态发生蛋白家族参与糖尿病诸多并发症的发生发展[14],如促进视网膜病变、心血管病变和糖尿病肾病等并发症的发展[15-17]。BMP4在炎症、肿瘤和其他消化道疾病发生发展的过程中,表达和分布发生特定的变化[2,5],在多种消化道及其他组织的肿瘤相关研究中发现BMP4具有促进肿瘤发生及侵袭转移的潜能,如肝细胞肝癌中高表达BMP4增强了肿瘤的侵袭性而其在胰腺癌中却表现出双相作用的特点[8,18]。在消化道疾病中,肠功能紊乱、内环境平衡破坏时,诸如炎症反应、动力紊乱及肿瘤发生等病理状态,异常表达的BMP4对于平滑肌功能可能存在抑制性调控作用,提示其参与胃肠稳态和动力的维持。本研究结果提示,BMP4及其受体BMPR1B在不同发育阶段大鼠的胃体、十二指肠和结肠中,广泛表达且表达水平和分布呈现一定的时空差异性。在胚胎消化道形成过程中,骨形态发生蛋白作为重要调节因素参与器官分化形成,在胚胎发育不同阶段,BMP4蛋白呈现差异性表达,且参与原肠组织分化与消化道各节段形成。在小鼠胚胎早期,诸如WNTs、FGFs、NOTCH、Hedgehog通路和BMP信号通路等均参与到早期消化道形成过程中[19-22]。在这其中,BMP信号通路直接调节或者间接与其他信号通路作用,调控内胚层的分化[17]。而BMP信号异常会导致许多器官发育不健全,如慢性肾病、成骨不全、骨关节炎、肺动脉高压、遗传性出血性毛细血管扩张症、Barrett食管、幼年息肉病等。大鼠出生后早期不能独立进食,消化道功能尚未完全成熟,随着3周龄大鼠能够独立进食,这种摄食行为的改变促使其胃体、十二指肠及结肠的功能逐渐完善成熟,与3周龄及8周龄大鼠十二指肠及结肠BMP4蛋白表达相较于5日龄大鼠差异性表达可能相关。这进一步提示在大鼠出生后BMP4的时间差异性表达可能参与胃体、十二指肠及结肠组织功能维持及分化过程。
形态学结果提示,BMP4蛋白主要由消化道上皮及腺体细胞分泌,其受体BMPR1B主要在黏膜层,消化道肌层散在分布。BMP4的这种分泌特点决定其不仅作用于分泌细胞本身,也对临近细胞及平滑肌细胞产生影响。在胃体免疫荧光双标染色可见,BMPR1B在黏膜层大量分布,肌层散在分布。在结肠升段中,BMP4受体主要分布在黏膜层,平滑肌层散在分布。在胚胎肠道发育阶段,BMP4协同Hedghog通路相关蛋白参与消化道环形肌和纵行肌分化,而在出生后不同阶段的差异表达和分布特点提示BMP4参与胃体及结肠平滑肌正常功能的维持。
综上,BMP4蛋白除在胚胎发育阶段对消化道形成分化起调控作用,在大鼠出生后肠道功能完善及维持的过程中,BMP4可能也起到重要作用。BMP4不仅参与正常组织细胞功能维持和分化,也参与消化道疾病的发生发展调控,因此对其在不同发育阶段大鼠消化道中的分布表达研究具有一定的临床提示意义。本研究通过研究BMP4及其受体在大鼠不同发育阶段胃肠道分布表达特点并结合既往研究结果,为研究胃肠炎性疾病以及胃肠道肿瘤提供新的潜在靶点。
