lncRNA CASC9通过靶向miR-195-5p/CDK6轴调控肺鳞癌的进展
2021-02-10张星星常丽花马运峰
张星星,杨 磊,刘 芳,张 瑾,常丽花,马运峰
(1西安交通大学基础医学院病原微生物与免疫学系,西安 710049;2西安医学院第二附属医院检验科;*通讯作者,E-mail:mayunfeng@xjtu.edu.cn)
肺癌是造成世界范围内癌症相关死亡的主要原因之一。肺癌主要分为小细胞肺癌和非小细胞肺癌。其中,非小细胞肺癌是肺癌的主要亚型,约占肺癌总数的85%[1]。根据组织学特性的不同,非小细胞肺癌又可分为肺腺癌、肺鳞状细胞癌(肺鳞癌)和大细胞癌[2]。据统计,约30%的非小细胞肺癌是肺鳞癌[3]。由于科研和医疗技术水平的提升,肺鳞癌的治疗取得了很大的进步,但肺鳞癌患者的预后依然很差[4]。因此,寻找肺鳞癌相关的潜在生物标记物并研究其分子机制对肺鳞癌诊断具有重要意义,同时可开发针对生物标记物的靶向治疗方法。
长链非编码RNA(lncRNA)作为RNA的类型之一,其长度超过200 nt,但不具备蛋白编码能力[5]。近年来,由于lncRNA在不同类型的人类疾病中发挥重要的调节作用而被广泛研究。研究发现lncRNA可通过调控基因的表达来影响肿瘤细胞的增殖、凋亡、迁移和侵袭等过程,进而影响肿瘤的进展[6-8]。尽管有报道称lncRNA CASC9(癌症易感性候选者9)与肺鳞癌的进展呈正相关[9],但其在肺鳞癌中的潜在分子机制还有待研究。
因此,本研究旨在揭示肺鳞癌中lncRNA CASC9的表达水平,并进一步探讨其潜在的分子机制。
1 材料与方法
1.1 主要材料
人胚肾细胞系293T、人正常肺细胞系BEAS-2B和肺鳞癌细胞系(SK-MES-1、NCI-H520、H1703和H2170)均购自ATCC细胞库。lncRNA CASC9的干扰物(si-CASC9-1和si-CASC9-2)和阴性对照(NC),以及lncRNA CASC9的过表达质粒(CASC9)和阴性对照(NC)购自上海Gene Pharma公司。miR-195-5p mimics和阴性对照NC mimics,以及miR-195-5p inhibitor和阴性对照NC inhibitor购自广州RiboBio公司。一抗(CDK6和GAPDH)和二抗购自美国CST公司。
1.2 细胞培养和转染
所有细胞放置在充满5% CO2的培养箱中,用含有10% FBS(HyClone, USA)、100 μg/ml链霉素和100 U/ml青霉素的DMEM培养基(HyClone, USA)培养。细胞接种于6孔板中,待融合度达到80%时,根据试剂商提供的说明书用lipofectamine 3000(Invitrogen,USA)进行转染,所有相关的转染分组在后续实验方法中根据实验目的进行分组。转染48 h后进行相关分析。
1.3 CCK8实验检测细胞增殖能力
为了研究CASC9对肺鳞癌细胞增殖能力的影响,首先将si-CASC9-1和si-CASC9-2转染到SK-MES-1和H1703细胞以干扰CASC9的表达。每种细胞中分3组:NC、si-CASC9-1和si-CASC9-2。
为了探讨CASC9和miR-195-5p是否通过CDK6影响癌细胞的增殖能力,用SK-MES-1和H1703细胞分别进行了如下2个实验的共转染:miR-195-5p与CDK6共转染实验中的分为NC inhibitor+si-NC组、NC inhibitor+si-CDK6组和miR-195-5p inhibitor+si-CDK6组;CASC9与CDK6共转染实验中分为NC+si-NC组、NC+si-CDK6组和CASC9+si-CDK6组。
根据以上不同的实验和分组进行铺板,肺鳞癌细胞以3 000/孔的数量接种在96孔板中,细胞转染0,24,48,72 h每孔加入10 μl CCK8(Dojindo)工作液,置于细胞培养箱中继续培养2 h,培养结束测定吸光度在450 nm处的吸光度值,细胞增殖能力根据CCK8试剂商提供的说明书计算。
1.4 Transwell实验检测细胞迁移和侵袭能力
利用Transwell实验去研究干扰CASC9对细胞迁移和侵袭能力的影响。SK-MES-1和H1703细胞经过转染后进行Transwell实验,每种细胞的转染分为NC组、si-CASC9-1组和si-CASC9-2组。转染48 h后收集细胞并重悬,取200 μl用无血清培养基重悬的(5×104个细胞)细胞悬液加到Transwell小室的上室中(8 μm pore, Corning, USA),下室加600 μl含有10%FBS的DMEM培养基。48 h后用4%的多聚甲醛室温固定15 min,以0.1%结晶紫染色15 min并用PBS冲洗干净。细胞侵袭实验中Transwell小室用基质胶(Corning, USA)进行包被,其余实验步骤同细胞迁移实验。在显微镜下随机选取5个视野进行拍照并计数。
1.5 荧光素酶报告基因实验验证CASC9和miR-195-5p的潜在靶基因
为了验证CASC9与靶基因miR-195-5p、以及miR-195-5p与靶基因CDK6之间的相互结合作用,我们构建了具有与miR-195-5p结合位点的CASC9或CDK6野生型和突变型报告质粒。将具有miR-195-5p结合位点的CASC9和CDK6的野生型序列与突变序列分别克隆到荧光素酶报告载体pmirGLO(Promega, China)中,得到CASC9野生型(CASC9-WT)、CASC9突变型(CASC9-MUT)、CDK6野生型(CDK6-WT)和CDK6突变型(CDK6-MUT)报告质粒。
CASC9和miR-195-5p荧光素酶实验分为:CASC9-WT+NC mimics组、CASC9-WT+miR-195-5p mimics组、CASC9-MUT+NC mimics组和CASC9-MUT+miR-195-5p mimics组。CDK6和miR-195-5p荧光素酶实验分为:CDK6-WT+NC mimics组、CDK6-WT+miR-195-5p mimics组、CDK6-MUT+NC mimics组和CDK6-MUT+miR-195-5p mimics组。根据以上分组将相应的物质转染到人胚肾细胞系293T中。转染48 h后收集细胞并裂解,利用荧光素酶检测试剂盒(Promega)检测荧光活性。
1.6 qRT-PCR实验检测CASC9、miR-195-5p和CDK6的表达
qRT-PCR实验用来检测肺正常细胞系(BEAS-2B)以及肺鳞癌细胞系(SK-MES-1、NCI-H520、H1703和H2170)中CASC9、miR-195-5p和CDK6的表达。
为了确认CASC9的干扰效率,将肺鳞癌细胞分别转染NC、si-CASC9-1和si-CASC9-2,用qRT-PCR实验检测转染后肺鳞癌细胞中CASC9的表达水平。
为了研究CASC9和miR-195-5p的表达是否具有相互调节作用,通过对肺鳞癌细胞中过表达miR-195-5p或干扰CASC9后,用qRT-PCR实验检测细胞中CASC9和miR-195-5p的表达。
根据以上不同实验目的处理细胞,之后收集细胞并向细胞样品中加入Trizol后超声匀浆提取总的RNA并检测RNA浓度。取1 μg RNA用逆转录试剂盒(TaKaRa)将RNA逆转录成cDNA。逆转录体系为:5×RNA逆转录酶2 μl;RNA 1 μg;DEPC水补到10 μl。反应程序为:42 ℃ 15 min,85 ℃ 5 s。之后通过SYBR Premix EX Taq试剂盒(TaKaRa)在荧光定量PCR仪中进行qRT-PCR反应。qRT-PCR反应体系为:2×Real time-PCR聚合酶10 μl;cDNA 0.5 μl;正向引物0.5 μl;反向引物0.5 μl,DEPC水补到20 μl。96孔板每孔加20 μl上述混合物后进行上机反应。反应程序为:95 ℃ 30 s;95 ℃ 5 s,60 ℃ 30 s(40个循环)。GAPDH作为CASC9和CDK6的内参,U6作为miR-195-5p的内参。
1.7 Western blotting实验检测CDK6的蛋白表达水平
为了确定miR-195-5p和CASC9对CDK6蛋白表达的影响,将肺鳞癌细胞分别转染NC mimics或miR-195-5p mimics、以及NC或CASC9。根据以上分组进行细胞转染。收集不同处理组的肺鳞癌细胞并提取蛋白。用RIPA裂解液(碧云天,北京)重悬细胞后将细胞裂解,4 ℃条件下以12 000 r/min离心10 min,收集蛋白上清液,用BCA蛋白定量试剂盒(碧云天,北京)检测蛋白浓度,并将蛋白样品煮沸变性。用10%的SDS-PAGE胶电泳分离蛋白质,电泳结束后将SDS-PAGE胶上的蛋白转移至PVDF膜。利用5%的脱脂奶粉将膜室温封闭2 h,随后将膜与一抗(CDK6,1 ∶1 000;GAPDH,1 ∶10 000)在4 ℃过夜敷育。TBST洗去未结合的一抗后使膜与辣根过氧化物酶(HRP)标记的二抗(1 ∶2 000)室温结合1 h。TBST洗3次,每次10 min。滴加ECL反应液(Thermo Fisher,USA)于膜上,室温进行显色反应,用Bio-Rad成像系统拍照。Image J软件进行灰度分析并计算相关蛋白的表达水平。
1.8 生物信息学分析
为了研究CASC9、miR-195-5p和CDK6在LUSC中的表达,以及预测CASC9和miR-195-5p的靶基因,我们采用生物信息学进行了相关分析。
利用GEPIA数据库分析了CASC9在癌组织和癌旁正常组织中的表达。在GEPIA数据库中输入“CASC9”并选择肺鳞癌“LUSC”,则可获得CASC9在癌组织和癌旁正常组织中表达的箱线图。
CASC9在肺腺癌患者不同阶段的表达利用Lnc2Cancer 3.0数据库完成,通过在Lnc2Cancer 3.0数据库中输入“CASC9”并选择肺鳞癌“LUSC”则可获得CASC9在肺腺癌患者不同阶段的表达。
利用lncbase和starbase数据库预测CASC9潜在的下游靶基因miRNAs。
通过在pubmed中搜索癌症与CASC9的4个靶基因相关的文献,发现miR-195-5p可作为肿瘤抑制因子,因此选择miR-195-5p作为CASC9的靶基因进行研究。
利用miR-pathway数据库分析miR-195-5p在包括肺腺癌等多种癌症中的表达。
miR-195-5p的靶基因由miRDB、miRWalk、TargetScan、starbase和miRTarbase数据库预测。
miR-195-5p的靶基因由京都基因与基因组百科全书(Kyoto Encyclopedia of Genes and Genomes,KEGG)数据库进行了通路富集分析。
利用GEPIA和UALCAN数据库分析CDK6在癌组织和癌旁正常组织中的表达。
1.9 统计学分析
本研究中所有数据均利用SPSS 21.0软件进行统计学分析。所有数据以平均值±标准差表示。两组间的比较采用独立样本t检验,多组间的比较采用单因素单方差分析。P<0.05认为差异有统计学意义。
2 结果
2.1 lncRNA CASC9在肺鳞癌中高表达
我们首先通过GEPIA数据库分析了CASC9在癌组织和癌旁正常组织中的表达,结果表明与癌旁正常组织相比,肺鳞癌癌组织中CASC9的表达明显升高(见图1A)。通过lnc2cancer数据库进一步分析发现CASC9的表达与肺鳞癌患者的肿瘤分期呈正相关(见图1B)。此外,我们利用qRT-PCR检测了人正常肺细胞和肺鳞癌细胞系中CASC9的表达量,结果表明CASC9在癌细胞的表达高于正常细胞(见图1C)。以上结果表明CASC9在肺鳞癌中高表达,可能与肺鳞癌的进展相关。
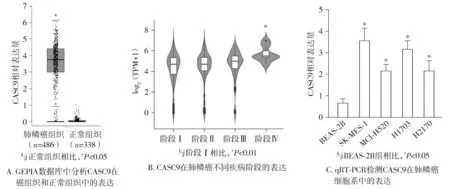
图1 CASC9在肺鳞癌癌组织和细胞中高表达Figure 1 CASC9 is high expressed in lung squamous cell carcinoma tissues and cells
2.2 干扰CASC9对肺鳞癌细胞增殖、迁移和侵袭能力的影响
qRT-PCR实验检测结果发现,肺鳞癌细胞中CASC9的表达在转染了si-CASC9后明显降低(见图2)。
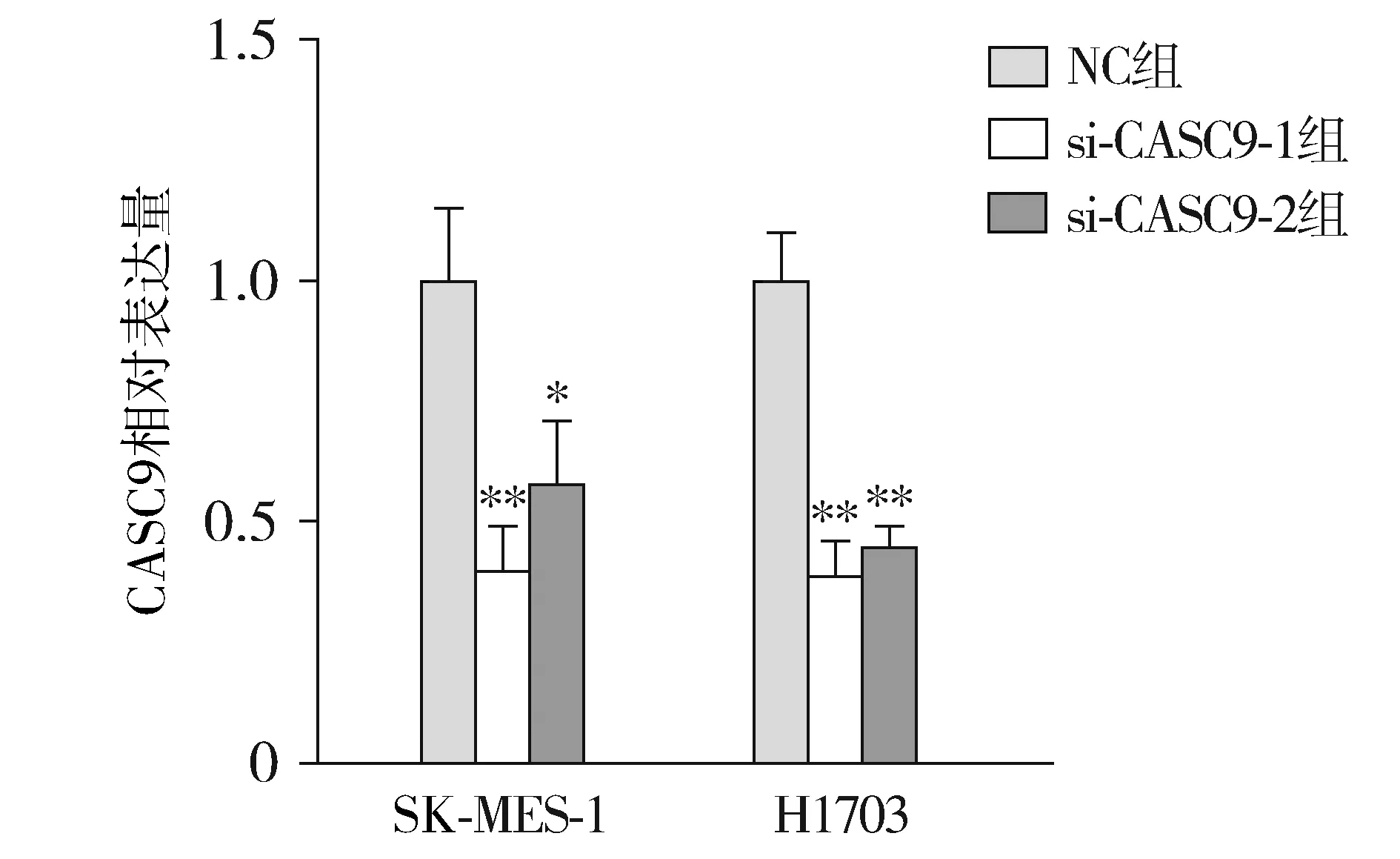
与NC组相比,*P<0.05,**P<0.01图2 CASC9的干扰效率Figure 2 The interference efficiency of CASC9
CCK8实验分析发现,肺鳞癌细胞中干扰CASC9后,细胞增殖能力显著降低(见图3)。
Transwell实验表明干扰CASC9后肺鳞癌细胞的迁移和侵袭能力受到显著的抑制(见图4)。
2.3 CASC9直接靶向miR-195-5p并调控其表达
为了研究CASC9促进肺鳞癌进展的分子机制,我们利用lncbase和starbase数据库预测CASC9潜在的下游靶基因miRNAs,在2个数据库中发现4个共同存在的潜在靶基因:miR-424-5p,miR-15a-5p,miR-195-5p,miR-15b-5p(见图5A)。通过对4个miRNAs在癌症中作用的检索发现miR-195-5p在多种癌症中发挥肿瘤抑制因子的作用,因此选择miR-195-5p作为CASC9的潜在靶基因来进一步研究。图5B为CASC9和miR-195-5p的潜在结合位点以及结合位点的突变序列。同时,利用miR-pathway数据库分析发现,miR-195-5p在NSCLS的两个重要亚型肺鳞癌和肺腺癌中的表达明显低于正常对照(见图5C)。
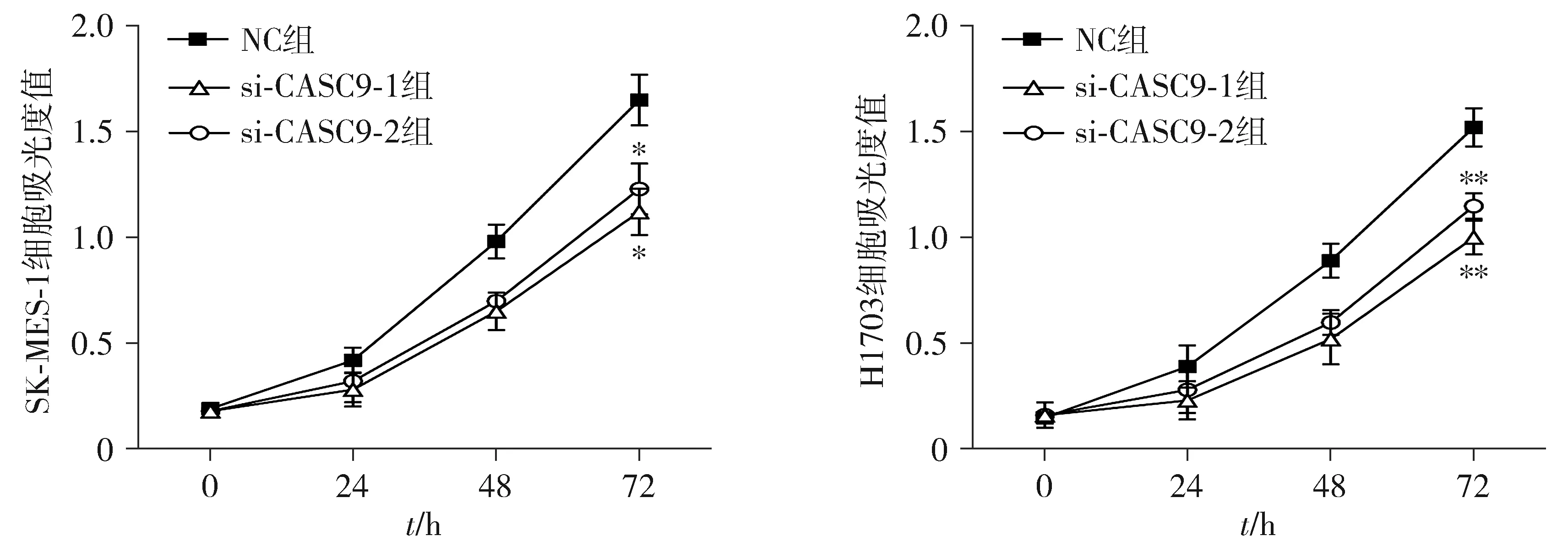
与NC组相比,*P<0.05,**P<0.01图3 CASC9对肺鳞癌细胞增殖的影响Figure 3 The effect of CASC9 on the proliferation of LUAD cells
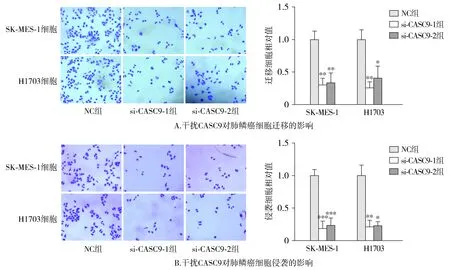
与NC组相比,*P<0.05,**P<0.01,***P<0.001图4 干扰CASC9对肺鳞癌细胞迁移和侵袭的影响Figure 4 The effect of CASC9 knockdown on the migration and invasion of LUAD cells
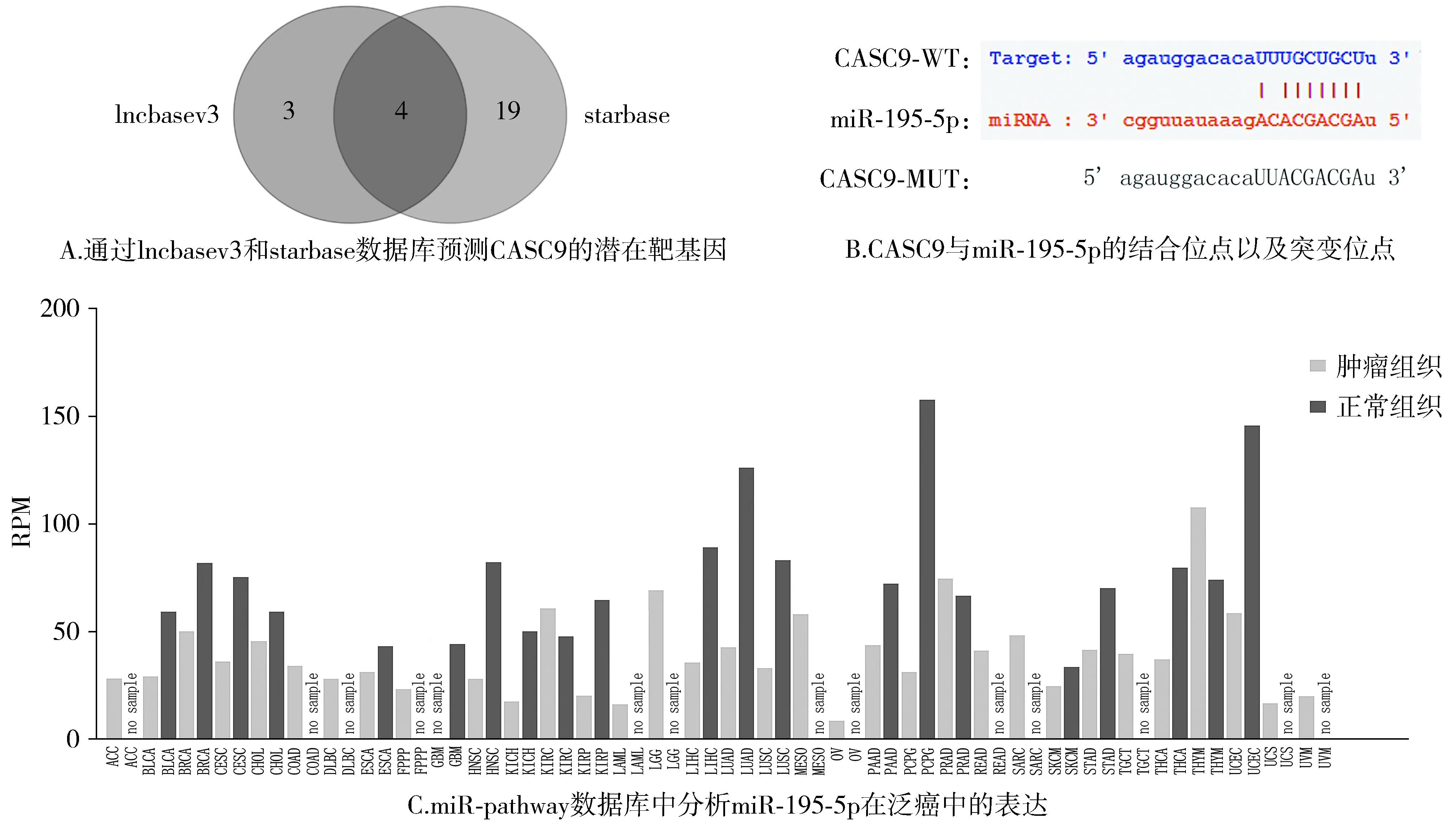
图5 CASC9靶向miR-195-5pFigure 5 CASC9 targets miR-195-5p
我们通过qRT-PCR进一步发现在肺鳞癌细胞系中miR-195-5p呈低表达(见图6A),表明miR-195-5p可能发挥抑制肺鳞癌的作用。为了证实CASC9与miR-195-5p之间的靶向关系,我们将具有与miR-195-5p结合位点以及结合位点突变的CASC9报告质粒(WT和MUT)与miR-195-5p mimics进行共转染,结果发现WT报告质粒处理组中荧光素酶活性显著低于MUT组(见图6B),表明CASC9可直接靶向miR-195-5p。通过对肺鳞癌细胞分别转染miR-195-5p mimics和si-CASC9,并利用qRT-PCR分析发现miR-195-5p和CASC9呈负向调控关系(见图6C,D)。因此,上述结果表明肺鳞癌中CASC9可直接靶向miR-195-5p并负调控其表达。
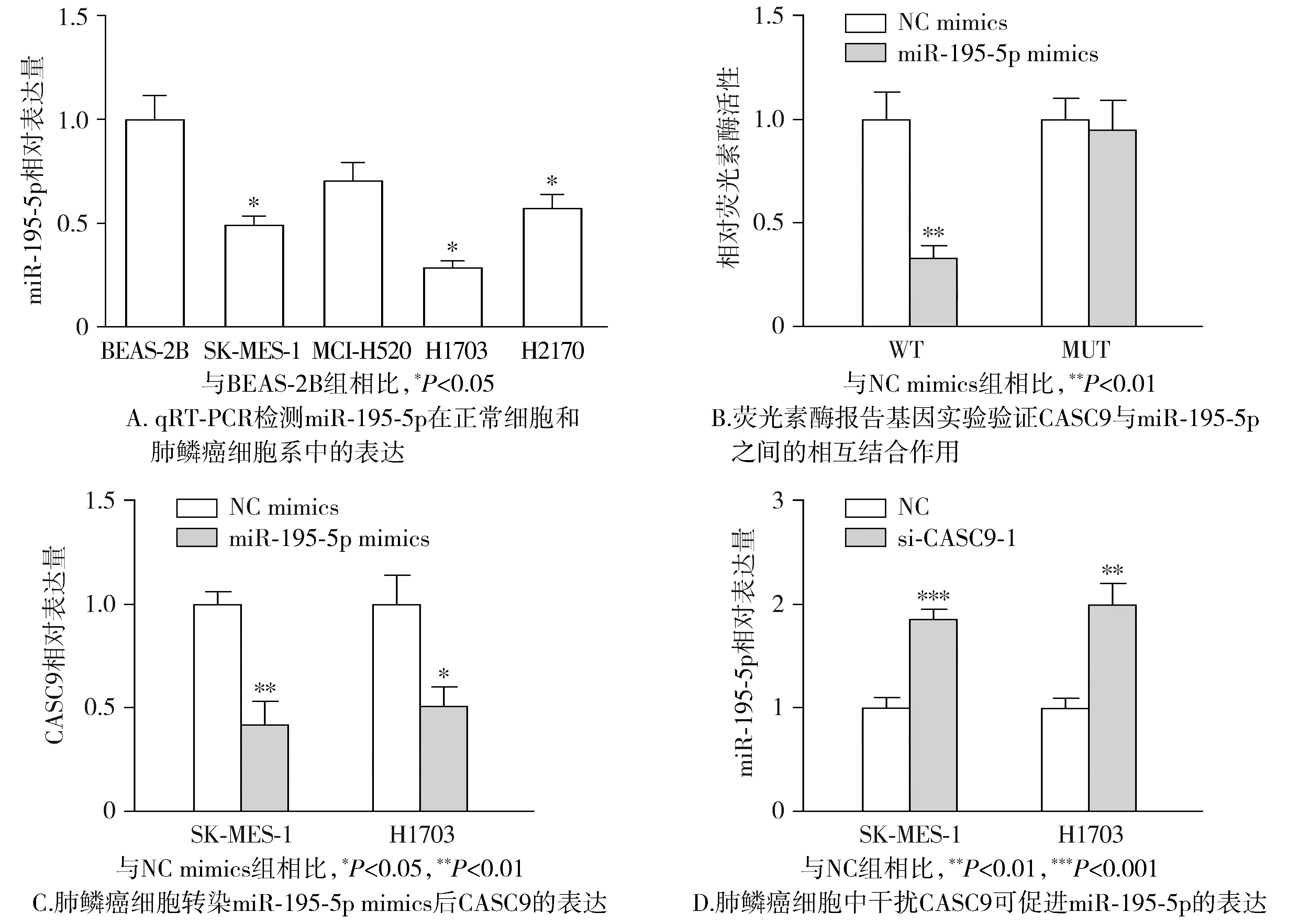
图6 CASC9靶向并调控miR-195-5p的表达Figure 6 CASC9 targets and regulates the expression of miR-195-5p
2.4 miR-195-5p靶向CDK6
我们通过miRDB、miRWalk、TargetScan、starbase和miRTarbase数据库预测了miR-195-5p的潜在靶基因,发现23个靶基因共同出现在5个数据库中(见图7A)。我们将此23个靶基因进行KEGG通路富集分析,发现CDK6与非小细胞肺癌相关(见图7B)。通过对GEPIA和UALCAN数据库的分析发现,CDK6在肺鳞癌癌组织中明显高表达(见图7C,D)。此外,荧光素酶报告基因实验表明miR-195-5p mimics可明显降低CDK6-WT组细胞的荧光活性(见图7E)。
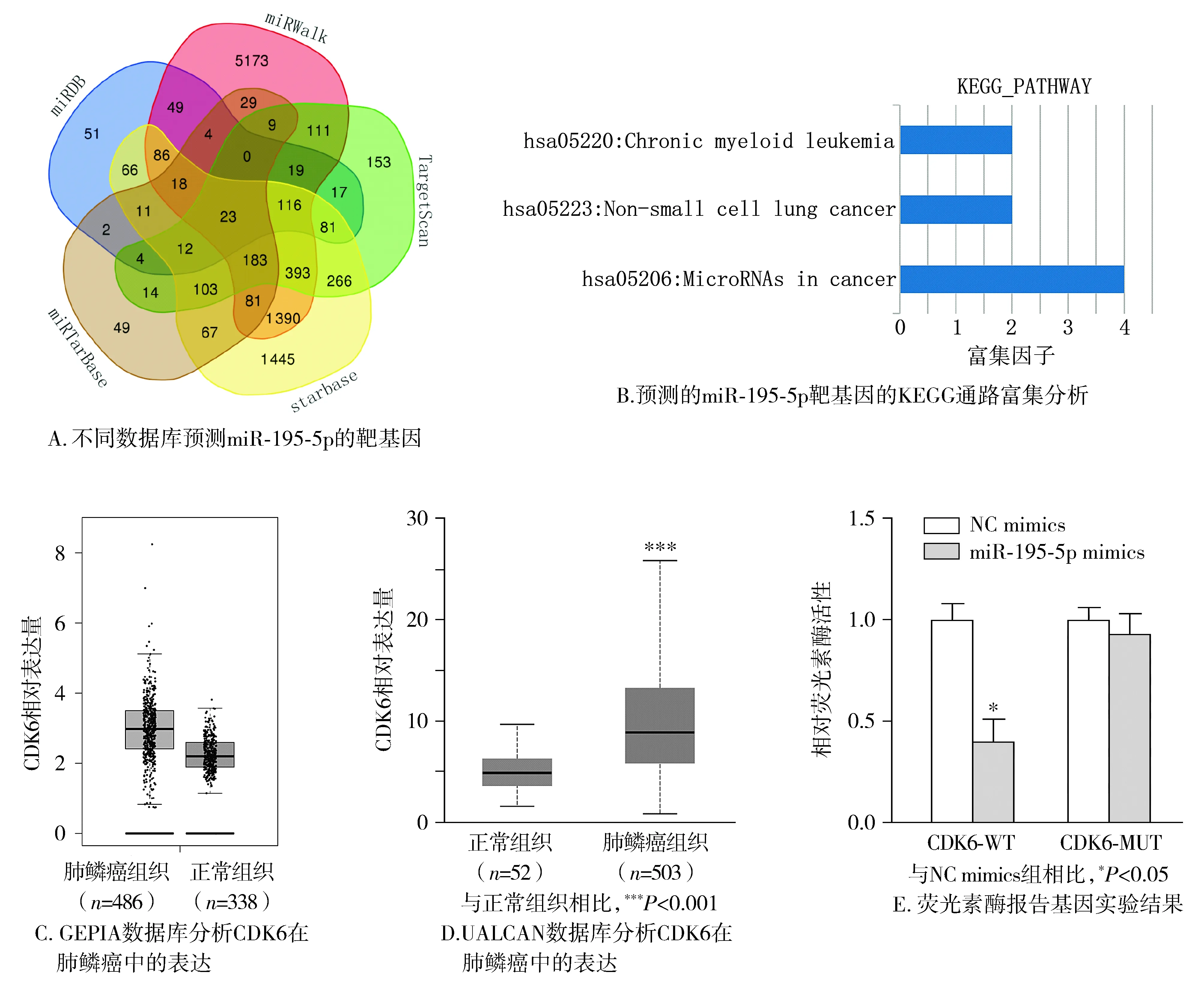
图7 miR-195-5p靶向CDK6Figure 7 miR-195-5p targets CDK6
2.5 CASC9/miR-195-5p调控CDK6的表达
qRT-PCR结果表明CDK6在肺鳞癌细胞中的表达高于正常细胞(见图8A)。随后,Western blotting结果发现过表达miR-195-5p后CDK6的表达显著被抑制(见图8B)。而肺鳞癌细胞中转染CASC9过表达质粒和阴性对照NC后,结果表明过表达CASC9可明显促进CDK6的表达(见图8C)。
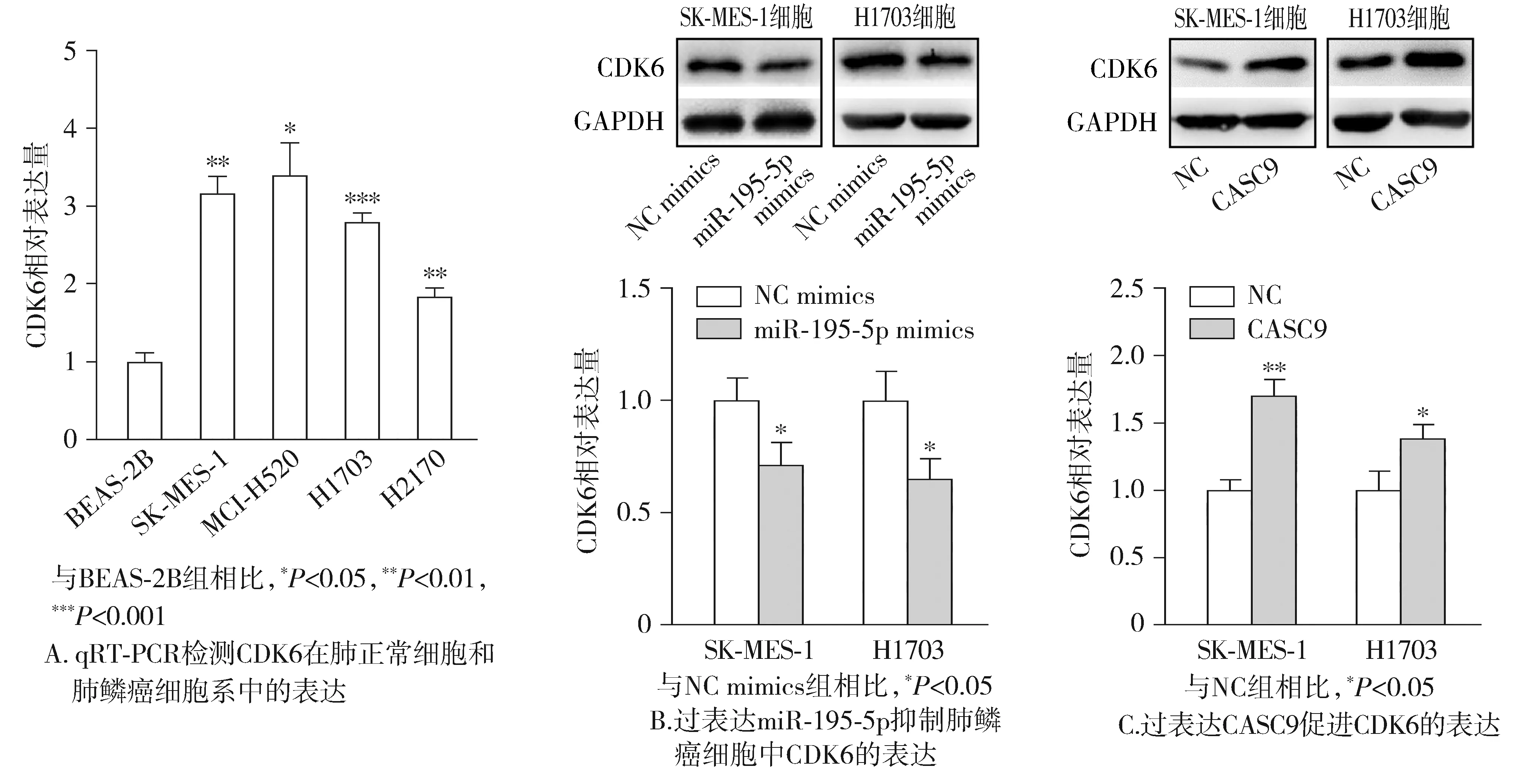
图8 CASC9/miR-195-5p调控CDK6的表达Figure 8 CASC9/miR-195-5p regulates the expression of CDK6
2.6 CASC9/miR-195-5p调控CDK6介导的细胞增殖
CCK-8结果发现干扰CDK6后细胞增殖能力明显减弱(见图9)。而营救实验表明,将miR-195-5p inhibitor或CASC9过表达质粒与si-CDK6共转染到肺鳞癌细胞后,敲低CDK6对细胞增殖能力的抑制作用显著被逆转(见图9)。这些结果表明CASC9/miR-195-5p通过介导CDK6调控癌细胞的增殖,从而促进肺鳞癌的进展。
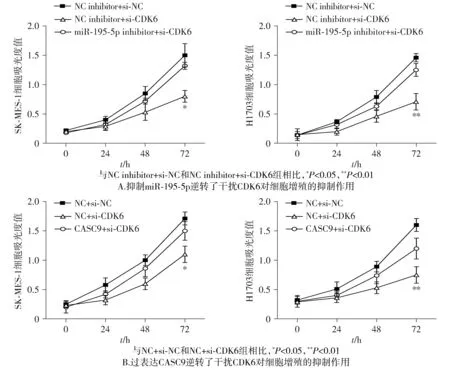
图9 CASC9/miR-195-5p调控CDK6介导的细胞增殖Figure 9 CASC9/miR-195-5p regulates CDK6-mediated cell proliferation
3 讨论
肺鳞癌作为常见的人类恶性肿瘤之一,目前对于其发病机制还未完全阐明。因此,有待进一步研究其内在致病相关的分子机制。据报道,lncRNA CASC9在多种癌症中通过发挥促癌基因的作用促进肿瘤的进展。例如,CASC9在结直肠癌中异常高表达,CASC9的高表达与患者的不良预后呈正相关,并且CASC9通过CPSF3-TGFβ2信号途径调控癌症的进展[10]。此外,最近的研究表明CASC9在膀胱癌中的表达显著上调,且与组织学分级、TNM分期和患者预后有关,沉默CASC9可抑制肿瘤的生长和膀胱癌的转移;而在分子机制上,CASC9通过正向调控FZD6的表达并激活Wnt/β-catenin信号传导途径,从而发挥促进膀胱癌的作用[11]。在本研究中,我们通过生物信息学分析和实验检测发现CASC9在肺鳞癌癌组织和细胞系中的表达显著上调,表明CASC9可能发挥促进肺鳞癌进展的作用。因此我们通过干扰CASC9来研究其对肺鳞癌细胞表型的影响。结果表明干扰CASC9可明显抑制癌细胞的增殖、迁移和侵袭能力。
lncRNA调控肿瘤进展的一种重要模式是通过充当竞争性内源性RNA(ceRNA)[12]。例如,LINC01133通过靶向miR-106a-3p调控APC表达和Wnt/β-catenin信号途径来抑制胃癌的进展[13]。在卵巢癌中,lncRNA PTAR可通过竞争性结合miR-101-3p上调ZEB1的表达,进而加速卵巢癌的EMT过程和转移的发生[14]。此外,lncRNA CDC6通过吸附microRNA-215而介导乳腺癌中CDC6的表达,因此促进乳腺癌的进展[15]。lnc00511在肺鳞癌中表达的升高与肿瘤分期相关,且通过抑制miR-150-5p和激活TADA1促进肺鳞癌的增殖和迁移[16]。为了研究CASC9在肺鳞癌中的作用机制,我们通过生物信息学的分析和荧光素酶报告基因实验发现miR-195-5p可作为CASC9的靶基因,其表达在肺鳞癌癌组织和细胞系中均降低,说明miR-195-5p在肺鳞癌中可能发挥抑癌的作用。同时,miR-195-5p与CASC9的表达呈负向调控关系。在本研究中,我们同样探讨了CASC9是否通过ceRNA的模式影响癌细胞的表型。我们首先通过生物信息学软件分析发现CDK6可能是miR-195-5p的潜在下游靶基因。已有研究发现CDK6在膀胱癌、结肠癌和卵巢癌[17-19]等多种癌症中高表达。我们进一步通过荧光素酶报告基因实验证明miR-195-5p可直接靶向CDK6。本研究发现,作为促癌基因,CDK6在肺鳞癌组织和细胞中的表达均升高。此外,我们发现过表达miR-195-5p可抑制CDK6的表达,而过表达CASC9可明显上调CDK6的表达。结合干扰CASC9后miR-195-5p表达升高的结果,本研究说明CASC9以ceRNA的模式影响了肺鳞癌的进展,即CASC9通过靶向miR-195-5p/CDK6信号轴调控肺鳞癌的进展。
总之,我们的研究表明CASC9在肺鳞癌中呈高表达,且CASC9的表达与肺鳞癌患者的肿瘤分级呈正相关。分子机制研究表明CASC9通过靶向抑制miR-195-5p上调癌基因CDK6的表达。因此,本研究结果表明CASC9/miR-195-5p/CDK6信号通路可能是肺鳞癌诊断或治疗的潜在分子靶点。
