伊犁马对补喂α-酮异己酸的吸收及其某些相关代谢的研究
2021-01-25赖晓鹤张文杰艾毕肯杨开伦
赖晓鹤,张文杰,韩 明,艾毕肯,刘 律,杨开伦
(新疆农业大学动物科学学院,新疆肉乳用草食动物营养重点实验室,新疆乌鲁木齐830052)
α-酮异己酸(KIC)是亮氨酸合成与分解的中间代谢产物,能够与亮氨酸通过支链氨基酸转氨酶(BCAT)进行可逆的转化,KIC也是肝脏生成β-羟基-β-甲基丁酸(HMB)的直接前体物质。研究表明,KIC具有促进肌肉蛋白质合成(Van和Edwarda,2005)、抑制蛋白质降解(Tischler 等,1982)、调节能量代谢(Moghei等,2016;Zhou 等,2010)及氮平衡(Sapir等,1983)等多种生理作用。在机体内KIC能够与亮氨酸相互转化,但KIC在降低血浆游离氨基酸浓度、促进骨骼肌蛋白质合成和降低血浆中尿素氮浓度等方面更加有效(Duan等,2019)。
Duan等(2019)给仔猪通过饲粮补喂1.25%KIC-Ca发现,其可促进肌肉蛋白质合成能力和抑制蛋白质降解。给禁食仔鸡补喂2.25 mg/kg的KIC能够起到抑制肌原纤维蛋白降解的作用(Nakashima等,2007)。研究发现,通过饲粮补喂1.25%的KIC-Ca能够促进仔猪肌肉组织中脂肪酸氧化,减少脂肪沉积(Duan等,2016)。桑丹等(2009)每天给绵羊分别补喂 0.5、1.0、1.5 g 的瘤胃保护性KIC-Ca发现,其能够调节氮平衡。每天补喂15 g的瘤胃保护性KIC-Ca能够增加羔羊体重(Kuhlman等,1988)。 目前,KIC作为营养添加剂的研究较少,将KIC作为一种添加剂补喂伊犁马,伊犁马对KIC的吸收及其对体内其他氨基酸会产生怎样的影响鲜见报道。本试验选用1岁伊犁马为研究对象,探究伊犁马对饲粮中补喂不同水平α-酮异己酸的吸收及其某些相关代谢情况,为KIC在马匹饲粮中的应用提供参考依据。
1 材料与方法
1.1 试验设计及饲粮组成 采取单因素多水平试验设计方法,挑选20匹年龄1岁,体重(295.25±25.15)kg的伊犁马,分成4组,每组 5匹,分别为对照组、试验Ⅰ组、试验Ⅱ组、试验Ⅲ组。试验饲粮由精料补充料和干草组成。不同组马匹所喂精料补充料不同。精料补充料配方及营养水平见表1,委托新疆伊犁州昭苏县骏力农牧科技有限公司生产加工。在试验饲粮的基础上,各试验组补喂不同水平的KIC,KIC以钙盐的形式进行补喂(购自张家港市思普生化有限公司,纯度为99%,有效浓度为87%)。玉米和次粉先按照1:1进行混匀,混匀后与KIC钙盐按照1:1制作成颗粒料,将颗粒料分为两份与精料补充料混合补喂。试验Ⅰ、Ⅱ、Ⅲ组每匹马KIC有效补喂量分别为6、12、18 g/d。本试验中KIC的补喂量参考了Yarrow等(2007)在运动员上的应用,选用低剂量0.02 g/kg的研究结果,进行为期25 d的补喂试验。精料补充料和干草中氨基酸含量见表2。

表1 精料补充料配方及营养水平(干物质基础)%
1.2 试验动物饲养管理 试验期间所有马匹均在同一马厩,单栏饲养。干草每天每匹马(8±0.8)kg, 分别在 08:30、13:30、18:30 和 23:30 分 4 次饲喂。精料补充料每天每匹马3 kg,分别于每天09:00和21:00分2次饲喂。先粗后精,先采食一定量干草,再补给精料补充料,待精料补充料采食完毕后,将马匹赶入活动圈,继续采食一定量的干草,自由饮水。每天按时打扫马厩,清除粪便,更换垫草。
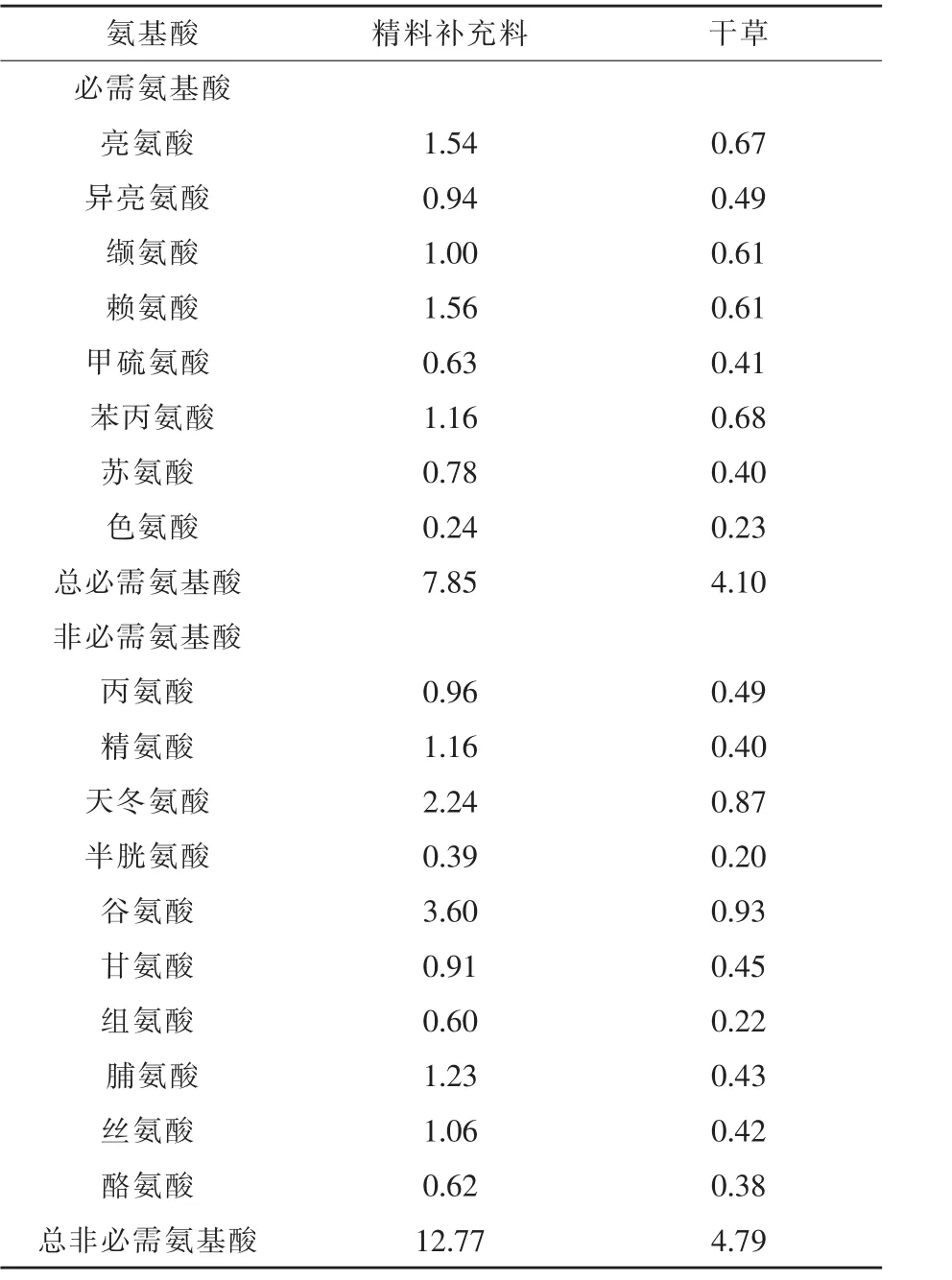
表2 精料补充料和干草中氨基酸含量(干物质基础)%
1.3 样品采集及指标的测定
1.3.1 血浆α-酮异己酸和β-羟基-β-甲基丁酸测定 在试验第25天采集血液,血液采集时间以补喂前 0 h 开始(8:30)以及补喂后的 1 h(10:00)、2 h(11:00)和 4 h(13:00),通过颈静脉采集血液置于肝素钠抗凝管中,以3500 r/min离心10 min,收集上清血浆,分装于1.5 mL Eppendorf管中,分装完毕后在-20℃冰箱中进行冻存。
血浆α-酮异己酸和β-羟基-β-甲基丁酸浓度均由北京普瑞华盛生物科技有限公司采用高效液相色谱串联质谱联用仪进行定量分析检测。
1.3.2 血浆游离氨基酸测定 在试验第25天采集血液,通过颈静脉于补喂后2 h(11:00)采集血液至肝素钠抗凝管中,以3500 r/min离心10 min,收集上清血浆,分装于1.5 mL Eppendorf管中,分装完毕后在-20℃冰箱中进行冻存。
血浆5-羟色胺浓度由北京华英生物技术研究所采用酶联免疫吸附法测定;血浆游离氨基酸浓度由北京普瑞华盛生物科技有限公司采用高效液相色谱串联质谱联用仪进行定量分析检测。
1.4 数据处理与分析 数据采用Excel 2010进行初步整理,试验结果以“平均值和标准误”表示。试验数据采用SPSS 19.0统计软件中的单因素方差分析(one-way ANOVA)进行差异显著性检验,差异显著用Duncan氏法进行多重比较。采用正交多项式对比,检验日粮中α-酮异己酸浓度增加的线性和二次效应。以P<0.05为差异显著的判断标准,P<0.01为差异极显著水平,0.05<P<0.10为有差异趋势。
2 结果与分析
2.1 伊犁马对α-酮异己酸的吸收 日粮α-酮异己酸补喂水平与伊犁马血浆中α-酮异己酸浓度的关系见图 1,当补喂 0 g/horse·meal(指每匹马每顿补喂的克数,下同)的KIC(对照组)时,血浆中KIC浓度最低;3 g/horse·meal(试验Ⅰ组)时,血浆中 KIC 浓度升高。且当 6 g/horse·meal(试验Ⅱ组)时,血浆中KIC浓度最高。随着KIC补喂水平的增加,血浆中KIC浓度的变化呈先上升后下降的趋势 (拟合方程为y=-0.0365x2+0.577+11.016,R2=0.9653)。结果显示,补喂不同水平的KIC对伊犁马血浆中KIC浓度无显著影响。
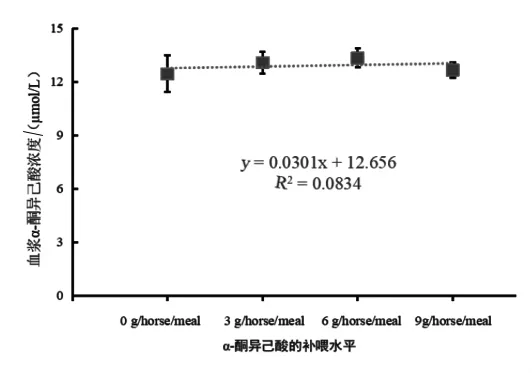
图1 日粮α-酮异己酸补喂水平与伊犁马血浆中α-酮异己酸浓度的关系
补喂α-酮异己酸对伊犁马血浆α-酮异己酸浓度的影响见表3。与对照组相比,补喂前(即0 h)各组间血浆KIC浓度差异不显著(P>0.05)。与对照组相比,补喂后1 h试验Ⅰ、Ⅱ组和Ⅲ组血浆KIC浓度分别提高了11.2%、7.86%和21.51%(P>0.05);补喂后2 h试验Ⅰ组、试验Ⅱ组和试验Ⅲ组血浆KIC浓度分别提高了8.66%、18.77%和8.84%,但均差异不显著(P>0.05);补喂后4 h各试验组均差异不显著(P>0.05)。3个试验组马匹血浆KIC浓度都在补喂后1 h出现峰值。
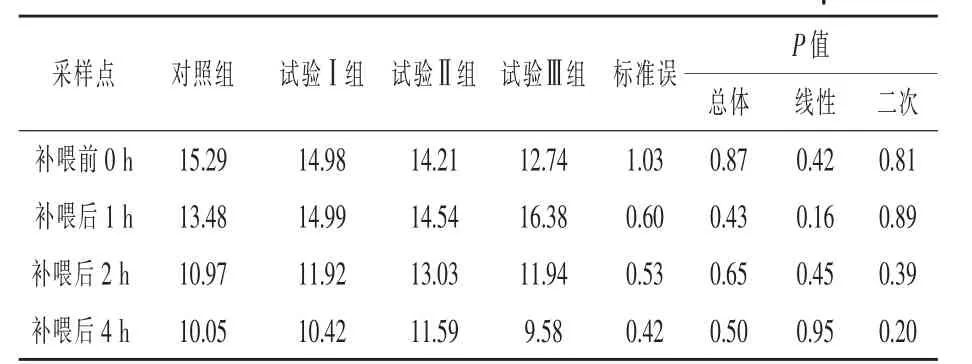
表3 补喂α-酮异己酸对伊犁马血浆中α-酮异己酸浓度的影响 μmol/L
2.2 补喂α-酮异己酸对伊犁马血浆β-羟基-β-甲基丁酸浓度的影响 日粮α-酮异己酸补喂水平与伊犁马血浆中β-羟基-β-甲基丁酸浓度的关系见图2,当KIC补喂水平为3 g/horse·meal(试验Ⅰ组)时,血浆中HMB浓度最低;且当6 g/horse·meal(试验Ⅱ组)时,血浆中HMB 浓度最高。随着KIC补喂水平的增加,血浆中HMB浓度的变化呈先下降后上升再下降的波浪趋势。结果显示,补喂不同水平的KIC对伊犁马血浆中HMB浓度无显著影响。

图2 日粮α-酮异己酸补喂水平与伊犁马血浆中β-羟基-β-甲基丁酸浓度的关系
补喂α-酮异己酸对伊犁马血浆β-羟基-β-甲基丁酸浓度的影响见表4。与对照组相比,补喂前0 h各组间血浆HMB浓度差异不显著 (P>0.05)。与对照组相比,补喂后1 h血浆HMB浓度各试验组均差异不显著(P>0.05);补喂后2 h试验Ⅰ、Ⅱ、Ⅲ组血浆HMB浓度分别提高了5.84%、15.74%和14.97%(P>0.05);补喂后4 h各试验组分别提高了8.95%、18.93%和14.58%,但均差异不显著(P > 0.05)。

表4 补喂α-酮异己酸对伊犁马血浆中β-羟基-β-甲基丁酸浓度的影响 μmol/L
2.3 补喂α-酮异己酸对伊犁马血浆游离氨基酸及其代谢物浓度的影响 补喂α-酮异己酸对伊犁马血浆游离氨基酸浓度的影响见表5。与对照组相比,各试验组血浆KIC浓度分别提高了8.66%、18.77%和8.84%,但均差异不显著(P>0.05);各试验组血浆HMB浓度均差异不显著 (P>0.05);试验Ⅱ组和试验Ⅲ组血浆亮氨酸浓度分别提高了22.40%和22.24%,且达到显著水平(P<0.05);试验Ⅰ组和试验Ⅲ组血浆异亮氨酸浓度降低了23.08%和32.34%,且达到极显著水平(P<0.01);试验Ⅰ组和试验Ⅲ组血浆缬氨酸浓度分别降低了15.75%和14.13%,且达到显著水平(P<0.05);试验Ⅲ组血浆赖氨酸、甲硫氨酸、精氨酸和谷氨酰胺浓度分别降低了 28.43%、22.22%、21.81%和23.60%,且均达到显著水平(P<0.05);各试验组血浆5-羟色胺浓度分别降低了7.5%、28.03%和10.28%,且试验Ⅱ组比试验Ⅰ组和试验Ⅲ组分别降低了22.22%和24.68%,组间差异均达到极显著水平(P<0.01);试验Ⅲ组血浆三甲基组氨酸浓度降低了18.74%,但差异不显著(P>0.05)。试验Ⅲ组血浆天冬酰胺浓度比试验Ⅱ组降低了26.50%,且组间差异达到显著水平(P<0.05),试验Ⅱ组血浆支链氨基酸浓度比试验Ⅰ组降低了18.80%,且达到显著水平(P<0.05)。
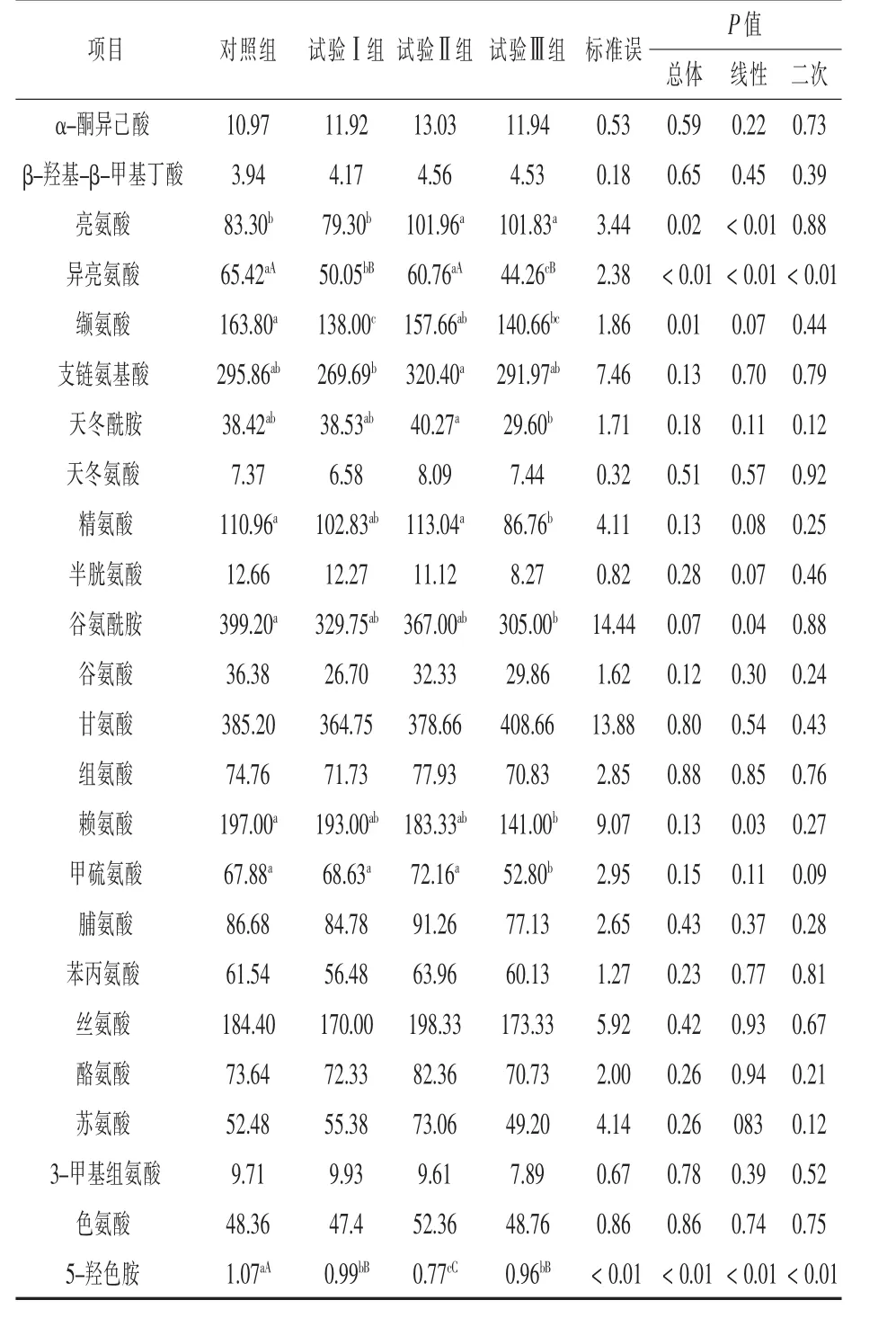
表5 补喂α-酮异己酸对伊犁马血浆氨基酸浓度的影响 μmol/L
3 讨论
3.1 伊犁马对α-酮异己酸的吸收 在动物机体内KIC有3种来源,从含有KIC的食物中直接摄取;也可以直接摄取KIC复合盐或通过摄取亮氨酸代谢产生KIC,进而提高血浆中KIC浓度。因此,本试验探究了日粮中补喂不同水平KIC与伊犁马血浆中KIC浓度的关系。本试验中,随着KIC补喂水平的增加,血浆中KIC浓度的变化呈先上升后下降的趋势。且当KIC补喂水平为6 g/horse·meal(试验Ⅱ组)时,血浆中KIC浓度最高。结果表明,补喂不同水平的KIC对伊犁马血浆中KIC浓度的影响不显著。KIC作为中间代谢物,在血浆中浓度处于相对稳定的状态。
测定动物饲喂前后血浆中间代谢物浓度的变化是动物营养消化代谢研究中常用的重要指标。从肝脏释放到血液中的KIC用于合成骨骼肌中的蛋白质或形成丙氨酸和谷氨酰胺。肝外组织中,KIC与亮氨酸可以相互转化,然后KIC可以重新合成亮氨酸或进一步氧化。本试验中,补喂KIC 1 h后血浆KIC浓度各试验组均达到了峰值。说明通过补喂KIC提高了伊犁马血浆中KIC浓度。本试验中,各试验组在补喂前及补喂后1、2、4 h血浆KIC浓度均呈先上升后下降趋势,KIC作为亮氨酸的中间代谢产物,在机体中的存留时间较短,通常会在肝脏组织中代谢或参与亮氨酸的相互转化。KIC的半衰期极短,血液中的KIC的代谢进行的很快(Giron等,2016),可能是因为KIC的氧化速率很快,这就解释了各组血浆中KIC的浓度在饲喂后1 h达到峰值,然后下降的原因。Duan等(2019)给仔猪补喂饲粮1.25%的KIC-Ca,1 h后血浆KIC浓度升高。Escobar等(2010) 发现给仔猪静脉滴注 400 μmol/kg·h 的KIC,1 h后血浆KIC浓度升高,与本试验结果一致。本试验中,对照组血浆中KIC浓度的变化呈下降趋势,各试验组血浆KIC浓度的变化表现为“单峰”模式(在补喂后1 h血浆KIC浓度达到峰值),且试验Ⅲ组血浆KIC浓度的变化表现为“单峰”模式最明显。
3.2 补喂α-酮异己酸对伊犁马血浆中β-羟基-β-甲基丁酸浓度的影响 机体内源性产生的 β-羟基-β-甲基丁酸(HMB)的量很少(Zhong等,2019)。KIC在肝细胞溶质中发生不可逆的氧化脱羧反应,大约5%的KIC通过KIC二氧化酶氧化生成HMB,生成的HMB大约有20%随尿液排出体外 (Molfino等,2013)。本试验中,随着KIC补喂水平的增加,血浆中HMB浓度的变化呈先下降后上升再下降的趋势,呈现出波浪状。值得注意的是,伊犁马中补喂低水平KIC(3 g/horse·meal)与未添加KIC相比,血浆中KIC浓度下降,可能是受到性别、个体差异和环境等外界因素的影响。当KIC补喂水平为6 g/horse·meal(试验Ⅱ组)时,血浆中HMB浓度最高。结果表明,补喂不同水平的KIC伊犁马血浆中HMB浓度的变化不显著。HMB作为KIC的代谢产物,通过内源性添加KIC,不会影响血浆中HMB浓度。
无论是内源性还是额外摄入的HMB,大约有30%的HMB被肌肉吸收 (Freund等,1978)。这就表明在高浓度时,HMB代谢的主要场所是肌肉组织(Van 和 Nissen,1992)。 研究表明,HMB不仅能够提高肌肉功能,还能提高脂肪代谢速率并降低蛋白质分解速率,促进组织细胞修复,减轻肌肉损伤并延缓肌肉疲劳(Norton和Layman,2006)。补喂KIC提高了血浆KIC浓度,从而导致血浆HMB浓度有所升高。本试验中,对照组血浆HMB浓度的变化呈下降趋势,试验I组和试验Ⅲ组在补喂前及补喂后1、2、4 h血浆HMB浓度的变化呈先上升后下降趋势,但试验Ⅱ组血浆中HMB浓度呈上升趋势。说明KIC在调节伊犁马血浆HMB浓度中可能存在剂量效应。Duan等(2019)给仔猪补喂1.25%KIC-Ca,1 h后血浆HMB浓度升高,与本试验结果一致。血浆KIC浓度的变化表现出的“单峰”模式,并没有使血浆HMB浓度出现相同的趋势,说明通过补喂KIC,机体代谢产生的HMB是十分有限的。因此补喂KIC在一定程度上增加了血浆HMB浓度,且试验II组血浆HMB浓度较高。
3.3 补喂α-酮异己酸对伊犁马血浆游离氨基酸及其代谢物浓度的影响
3.3.1 补喂α-酮异己酸对伊犁马血浆亮氨酸及支链氨基酸浓度的影响 亮氨酸作为必需氨基酸之一,必须通过日粮摄入的方式来维持机体正常生长及生产活动正常进行(Wu,2014)。本试验中,通过给伊犁马饲粮中补喂KIC可显著提高血浆中亮氨酸浓度,与对照组相比,试验Ⅱ组和试验Ⅲ组血浆亮氨酸浓度分别提高了22.40%和22.24%。血浆亮氨酸浓度升高,导致血浆缬氨酸和异亮氨酸浓度显著下降。由于三种氨基酸之间互为拮抗关系,亮氨酸增加必定会影响其他一种或两种氨基酸在机体内的吸收和利用 (Papet等,1988)。支链氨基酸对氨基酸与蛋白质的代谢具有调节作用,尤其是亮氨酸可以通过激活雷帕霉素靶蛋白(mTOR)信号通路调节骨骼肌的蛋白质合成(Wilkinson 等,2013;Suryawan 等,2011),而异亮氨酸和缬氨酸可以保证三种支链氨基酸之间的平衡(Suryawan 等,2013)。 Duan 等(2019)给仔猪补喂1.25%的KIC-Ca提高了血浆亮氨酸浓度,血浆缬氨酸、异亮氨酸浓度降低,与本试验结果一致。本试验中,试验Ⅰ组与试验Ⅱ组相比,血浆中支链氨基酸浓度显著下降。由于马匹血浆中游离氨基酸浓度会受到性别、环境和温度等因素影响 (Hackl等,2006),因此补喂不同水平的KIC对血浆支链氨基酸浓度的影响有待进一步研究。
3.3.2 补喂α-酮异己酸对伊犁马血浆3-甲基组氨酸浓度的影响 3-甲基组氨酸是组氨酸通过蛋白质合成甲基化后生成的氨基酸,主要存在于机体内骨骼肌蛋白的肌球蛋白和肌纤蛋白中。骨骼肌中蛋白质周转主要包括:骨骼肌收缩蛋白、肌动蛋白、肌球蛋白的周转,会导致血浆中3-甲基组氨酸释放,3-甲基组氨酸不能被机体重新利用合成蛋白,最终被排出体外(Rathmacher和Nissen,1998)。在一定程度上3-甲基组氨酸也可作为肌原纤维蛋白水解的指标(桑丹等,2009)。本试验中,血浆中3-甲基组氨酸浓度无显著变化,但整体呈下降趋势,且试验Ⅲ组血浆3-甲基组氨酸浓度降低了18.74%。研究发现,给禁食的雏鸡补喂2.25 mg/kg的KIC降低了血浆3-甲基组氨酸浓度(Nakashima 等,2007),Sapir等(1983)给术后患者每天静脉滴注70 mmol的KIC-Na降低了血浆3-甲基组氨酸浓度,与本试验结果相似。由于本试验中KIC的补喂量分早晚两次补喂,因此,伊犁马血浆中3-甲基组氨酸浓度的变化可能与KIC的补喂量有关。本试验中没有收集马尿,进而得到3-甲基组氨酸的排出量,因而没有直接证据表明动物机体内蛋白质的降解情况。但从本试验中的马匹增重情况可以得出,与对照组相比,各试验组的平均日增重有升高趋势。因此,KIC可能有提高伊犁马增重的趋势。
3.3.3 补喂α-酮异己酸对伊犁马血浆其他氨基酸及其代谢物浓度的影响 血浆游离氨基酸浓度反映了动物摄入饲粮氨基酸的水平。谷氨酰胺是马驹血浆中浓度最多的氨基酸 (Manso等,2009)。本试验中,试验Ⅲ组降低了血浆谷氨酰胺、赖氨酸、甲硫氨酸浓度。说明补喂KIC可能会影响伊犁马对某些氨基酸的吸收。赖氨酸和甲硫氨酸分别作为第一、第二限制性氨基酸,血浆赖氨酸和甲硫氨酸浓度降低,说明KIC维持了与限制性氨基酸的平衡,增加了伊犁马对限制性氨基酸的吸收利用。血浆精氨酸浓度降低,推测血浆中的精氨酸可能是参与了机体其他氨基酸的合成。研究发现,通过给仔猪静脉滴注400 μmol/kg·h的 KIC可降低血浆精氨酸、谷氨酰胺浓度(Escobar等,2010);Duan 等(2019)也证实了仔猪补喂1.25%的KIC-Ca可降低血浆中精氨酸浓度,与本试验结果一致。因此,KIC可能促进了机体蛋白质或氨基酸的利用率。脑中游离色氨酸浓度的变化与支链氨基酸浓度有关,因为支链氨基酸能竞争抑制色氨酸在血脑屏障上的载体——白蛋白通道(Leite等,2010)。色氨酸是5-羟色胺的前体物质,大脑吸收过多血液运输传递的游离色氨酸会引起脑内5-羟色胺增加,从而引起中枢疲劳(Davis等,2000)。本试验中血浆5-羟色胺浓度显著下降。说明KIC可能在抑制机体中枢疲劳方面具有积极作用。本试验中,各组血浆天冬氨酸、天冬酰胺和精氨酸浓度的变化呈下降趋势,但试验Ⅱ组相比其他各试验组血浆天冬氨酸、天冬酰胺和精氨酸浓度较高。关于补喂不同水平的KIC对氨基酸代谢的影响有待进一步研究。
4 结论
本试验结果表明,补喂α-酮异己酸提高了伊犁马血浆中α-酮异己酸浓度,且各试验组均在补喂后1 h达到峰值;提高了血浆亮氨酸浓度;降低了血浆中支链氨基酸和5-羟色胺的浓度。通过补喂不同水平的α-酮异己酸,改变了伊犁马对某些氨基酸的吸收利用。
