miR-200c 抑制自噬对肝癌耐药细胞HepG2/ADM化疗敏感性的影响*
2020-12-29陈有科高良辉黄小龙
陈有科 高良辉 符 誉 黄小龙
海南医学院第一附属医院肝胆胰外科 (海南 海口, 570102)
化疗是肝癌临床治疗的一个重要手段,但是化疗期间原发性或继发性肿瘤多药耐药(MDR)的产生常使化疗效果并不理想[1]。MDR的机制复杂多样,自噬是细胞对抗环境应激的一种保护性反应,近年来的研究发现自噬参与了肿瘤MDR的过程[2]。微小RNA(miRNA) 是体内一小段非编码核酸序列,其可以调控肿瘤的发生、发展、浸润、转移或耐药过程[3],微小RNA-200c(miR-200c) 属于miR-200家族成员,研究发现miR-200c水平是肝细胞癌的独立危险因素,参与肿瘤远处转移的调节,与患者的临床预后密切相关[4]。本研究以肝细胞癌HepG2细胞株,以及其多药耐药HepG2/ADM细胞株为对象,探讨miR-200c对在自噬的调控以及对肝癌MDR的影响。
1 材料与方法
1.1 主要试剂与仪器 人肝癌HepG2和HepG2/ADM细胞株购自广州吉赛生物科技有限公司,多柔比星购于美国Sigma公司,RIPA蛋白裂解液、细胞MTT测试试剂盒购自美国Promega公司,miR-200c(forward,5′-AGGGGTGAGACTAGGCAGGTTGG-3;reverse,5′-CCAGGTTGCAGTCCAAGCA-3′)和内参U6(forward,5′-CGCTTCGGCAGCACATATAC-3′;reverse,5′-TTCACGAATTTGCGTGTCAT-3′)引物由上海生工科技有限公司设计合成,RT-PCR逆转录试剂盒和SYBR Green qPCR superMix荧光定量试剂盒购自日本TaKaRa公司产品。脂质体2000和Trizol购自美国Invitrogen公司,含有miR-200c类似物序列(miR-200c mimics)和阴性对照无关序列(negative control miRNA)的pEZ-M02-UBQLN1质粒购自上海吉玛制药技术有限公司。Beclin-1、LC3Ⅱ和β-actin抗体购于美国Cell signal公司。酶联免疫检测仪购自美国Bio-Rad公司,实时PCR仪购自美国ABI公司,透射电镜购自东京杰尔有限公司,超薄切片机购自日本JEOL公司,细胞培养箱购自美国Thermo Fisher公司。
1.2 实验分组和细胞转染 肝癌和肝癌耐药细胞分为4组:HepG2组、HepG2/ADM组、阴性转染对照组和miR-200c类似物转染组。HepG2组和HepG2/ADM组不进行质粒转染,阴性转染对照组HepG2/ADM细胞转染对照质粒miRNA- pEZ-M02-UBQLN1质粒,miR-200c类似物转染组HepG2/ADM细胞转染miR-200c mimics- pEZ-M02-UBQLN1质粒,按照脂质体2 000试剂盒说明书操作,阴性转染对照组和miR-200c类似物转染组细胞换无血清的培养液,脂质体2 000处理细胞后加入相应质粒转染2 h,去除培养液,换用含10%胎牛血清的培养液。培养48 h后收集细胞,实时PCR方法检测转染效果,采用2-△△CT法计算miR-200c基因的相对表达量。
1.3 MTT方法测定细胞对化疗药物的敏感性 4组细胞分别消化为单细胞悬液,按2 000个/孔接种于96孔板,细胞贴壁后加入不同浓度的多柔比星(100、50、25、12.5、6.25、3.125、1.5625、0.78 125 μmol/L),培养48 h后每孔加入含10 μl MTT的培养基200 μl,培养箱中孵育4 h后去上清,加入100 μl DMSO溶解结晶,酶标仪测定490 nm处吸光度,计算多柔比星作用于各组细胞的IC50值。
1.4 透射电子显微镜(TEM)分析细胞自噬体 收集4组细胞,3 000 g离心10 min后用1% OsO4、0.1 mol/L碳酸钙缓冲液固定细胞4 h,梯度乙醇脱水后嵌入环氧丙烷,超薄切片机制作80 nm切片,1%醋酸铀酯和0.2%柠檬酸铅染色,TEM下观察自噬体,计算每个高倍视野下(8 000×)自噬体数。
1.5 自噬相关蛋白Beclin-1、LC3Ⅱ的表达 收集4组细胞提取细胞总蛋白,Western Blot方法检测自噬相关蛋白Beclin-1、LC3Ⅱ的表达,ImageJ软件对照内参分析目的蛋白的相对表达量。
1.6 统计学方法 用SPSS16.0软件进行分析,多组均数间比较采用方差分析,组间两两比较采用LSD-t检验,以P<0.05为差异有统计学意义。
2 结果
2.1 各组细胞miR-200c表达的差异及对化疗药物的敏感性(IC50)情况 见表1。
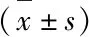
表1 各组细胞miR-200c表达及IC50值情况
2.2 miR-200c影响各组细胞自噬情况 HepG2组、HepG2/ADM组、阴性转染对照组和miR-200c类似物转染组高倍视野下自噬体分别为(0.16±0.04)个、(4.82±1.03)个、(4.77±1.01)个和(1.35±0.33)个,其中HepG2/ADM组自噬体显著多于HepG2组(P<0.01),HepG2/ADM组和阴性转染对照差异无统计学意义(P>0.05),miR-200c类似物转染组自噬体显著少于HepG2/ADM组(P<0.01),见图1。
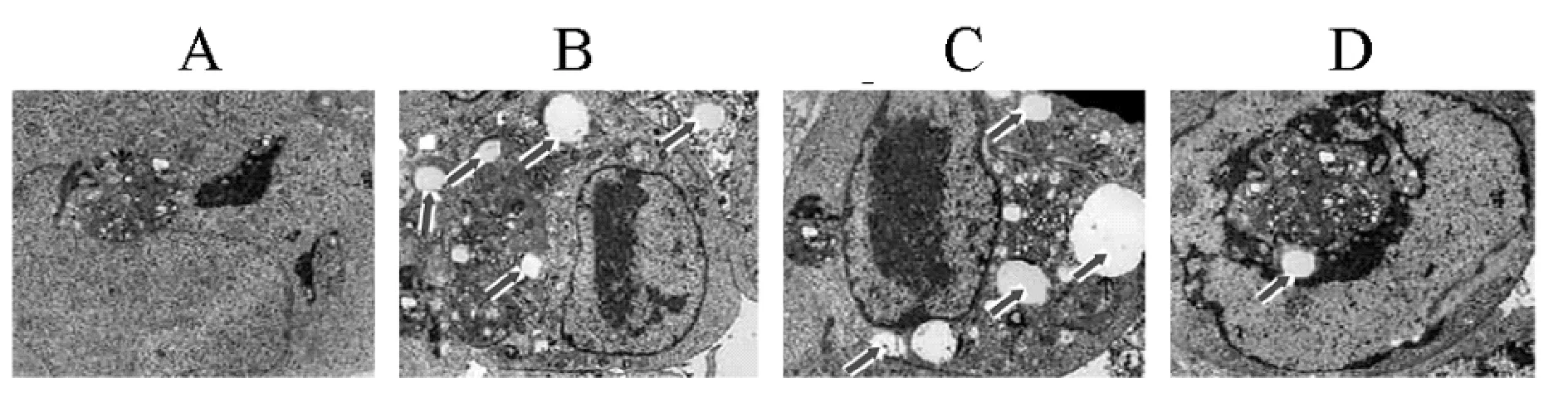
图1 各组细胞的自噬体 (TEM,8 000×)
箭头标示处为自噬体,A.HepG2组,B.HepG2/ADM组,C.阴性转染对照组,D.miR-200c类似物转染组
2.3 miR-200c对HepG2/ADM细胞自噬蛋白的影响 HepG2组、HepG2/ADM组、阴性转染对照组和miR-200c类似物转染组Beclin-1蛋白的相对表达量分别为(0.18±0.02)、(0.57±0.11)、(0.55±0.09)和(0.27±0.05),LC3-Ⅱ蛋白的相对表达量分别为(0.11±0.02)、(0.63±0.14)、(0.60±0.13)和(0.21±0.04),其中HepG2/ADM组Beclin-1和LC3-Ⅱ蛋白相对表达量显著高于HepG2组(P<0.01),HepG2/ADM组和阴性转染对照组间差异无统计学意义(P>0.05),miR-200c类似物转染组Beclin-1和LC3-Ⅱ蛋白相对表达量显著低于HepG2/ADM组(P<0.01),见图2。
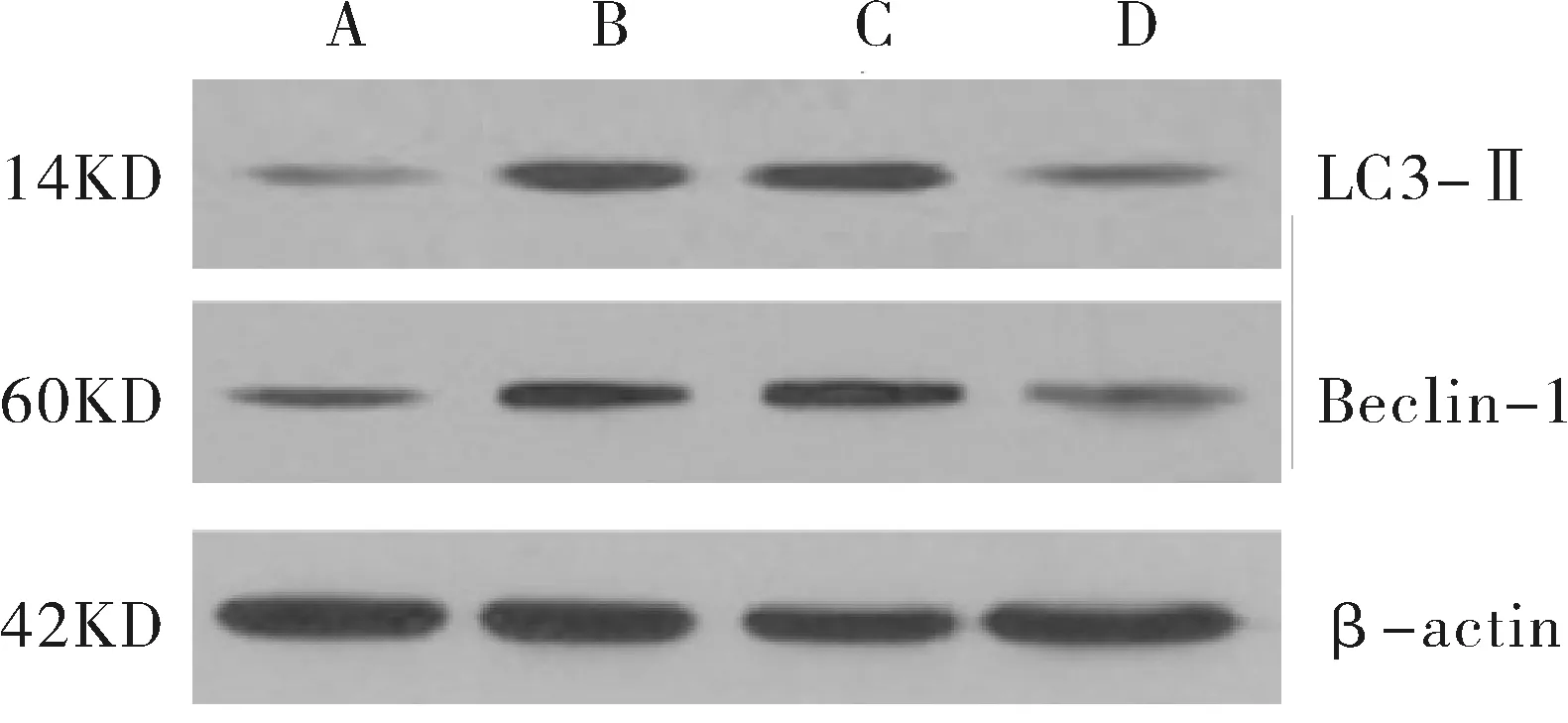
图2 Beclin-1和LC3-Ⅱ蛋白表达的情况A.HepG2组,B.HepG2/ADM组,C.阴性转染对照组,D.miR-200c类似物转染组
3 讨论
肿瘤的MDR机制极其复杂,有多种因素的参与,其中miRNA对靶基因的调节是一个重要的原因[5],miR-200c广泛存在于生物体,研究显示其在肝癌中可以发挥抑癌作用[6],Koo等[7]研究发现上调miR-200c可以增加肿瘤细胞对化疗的敏感性。本研究使用HepG2和HepG2/ADM细胞,结果显示多柔比星对HepG2/ADM细胞IC50值显著高于HepG2细胞,检验了HepG2/ADM细胞的耐药性,可以用于后续的研究,PCR分析显示HepG2/ADM细胞miR-200c的相对表达量显著低于HepG2细胞,说明了miR-200c低表达可能参与了肝癌的MDR。丁红等[8]的研究发现上调 miR-200c 表达可以靶向人肝癌干细胞,拮抗化疗耐药性,提示了靶向miR-200c可以调节肝癌的MDR。本研究进一步在HepG2/ADM细胞中转染miR-200c类似物过表达miR-200c,结果显示多柔比星对miR-200c类似物转染组细胞的IC50值显著低于HepG2/ADM组,也说明了上调miR-200c可以克服肝癌的MDR。
自噬是一种重要的细胞生存机制,在正常生理过程中起着“管家”的作用,清除受损的细胞器、抵御病原体、调节生长和衰老,自噬调节缺陷在许多疾病中起着重要作用,调节自噬也有助于癌症的治疗[9]。自噬也参与到肿瘤MDR的调节中,王浩等[10]抑制大肠癌细胞的自噬活性,发现细胞凋亡率增加,可以恢复部分药物敏感性,提示了调节细胞自噬可以作为一个抗肿瘤MDR的途径。Beclin-1和LC3-Ⅱ蛋白是自噬激活的标志蛋白[11],本研究发现HepG2/ADM组自噬体、Beclin-1和LC3-Ⅱ蛋白显著多于HepG2组,提示了肝癌耐药中存在自噬的过度激活。Sun等[12]发现miR-200c可以抑制乳腺癌细胞的自噬激活,从而显著增加细胞的放疗敏感性。本研究发现miR-200c类似物转染组自噬体、Beclin-1和LC3-Ⅱ蛋白显著低于HepG2/ADM组,说明了miR-200c可以抑制HepG2/ADM细胞的自噬激活,MTT细胞增殖分析也显示了miR-200c类似物转染组细胞对多柔比星的敏感性得到了显著改善。
综上所述,本研究显示上调miR-200c表达可以抑制HepG2/ADM细胞自噬,增加HepG2/ADM细胞对化疗药物的敏感性。
