miR-656-3p靶向ATP10A基因调控阿尔茨海默病模型细胞凋亡的分子机制
2020-12-09朱竹君祁英杰于明
朱竹君 祁英杰 于明
(江苏大学 1附属金坛医院神经内科,江苏 常州 213200;2附属医院神经内科)
阿尔茨海默病(AD)又称老年痴呆,其发生、发展及病理改变过程与环境、遗传因素等紧密相关〔1,2〕,但其病理演进过程的分子机制尚不明确。AD是一种神经退行性疾病,主要特征为认知功能障碍、记忆损害等〔3,4〕。微小RNAs(miRNA/miR)是一类长18~25 nt、广泛表达于哺乳动物中的非编码小RNA分子,具有高度保守性,含有与下游靶基因miR的3′非翻译区(UTR)互补的碱基序列,可抑制或降解靶基因miR,从而调控转录后水平。研究表明,miR异常表达与肿瘤、神经系统疾病等密切相关〔5,6〕。多种miR在AD演进过程中表达量异常,参与AD发生发展过程〔7,8〕。miR-656-3p在AD演进过程中表达量异常〔9〕,调控其发生发展过程,但miR-656-3p具体的作用机制尚不完全清楚。本研究旨在探讨miR-656-3p在AD中作用及相关机制,为进一步阐明AD发生发展的分子机制提供理论基础及临床新药的研发提供潜在分子靶点。
1 材料与方法
1.1实验主要试剂和仪器 SH-SY5Y细胞购自中国科学院上海细胞库;DMEM培养基、胎牛血清购自美国Gibco公司,青链霉素、细胞裂解液、双荧光报告基因检测试剂盒购自中国碧云天公司,胰酶、二喹啉甲酸(BCA)蛋白定量试剂盒、增强化学发光(ECL)试剂盒购自美国Thermo公司,Trizol试剂购自北京康为世纪生物科技有限公司,逆转录试剂盒、荧光定量聚合酶链反应(qPCR)试剂盒购自大连宝生物工程有限公司,噻唑蓝(MTT)试剂盒、凋亡试剂盒购自南京凯基生物科技有限公司,Lipofectamine2000购自美国Invitrogen公司,miR-656-3p 模拟物及阴性对照(NC)、anti-miR-432-5p、anti-NC及si-NC和si-ATP10A购自上海吉玛生物有限公司,聚偏氟乙烯(PVDF)膜购自美国Millipore公司,兔抗腺苷三磷酸酶(ATP)10A抗体,β-肌动蛋白(β-actin)抗体购自美国Santa Cruz Biotechnology公司,辣根过氧化物酶标记二抗购自武汉博士德生物科技有限公司,ATP10A野生型(ATP10A-wt)和ATP10A突变型(ATP10A-mut)荧光素酶报告载体购自广州锐博生物有限公司。细胞培养箱购自美国Thermo公司,超净工作台购自上海博翎有限公司,qPCR仪、PCR仪、酶标仪购自美国Bio-Rad公司。
1.2细胞的培养和分组 SH-SY5Y细胞培养于DMEM培养基,含有10%胎牛血清,放入37℃、5%CO2培养箱中,次日更换培养液,待显微镜下观察到SH-SY5Y细胞长满培养瓶底部,加入适量0.25%胰酶消化1~3 min,按1∶3比例传代培养。选取生长状态良好的SH-SY5Y细胞分为对照组、模型组、NC组、miR-656-3p组、anti-NC组、anti-miR-656-3p组、si-NC组、si-ATP10A组,对照组为正常培养的SH-SY5Y细胞,模型组为采用10 μmol/L的淀粉样蛋白(Aβ25~35)诱导SH-SY5Y细胞构建的AD模型细胞,NC组、miR-656-3p组为模型组细胞分别转染NC、miR-656-3p模拟物,anti-NC组、anti-miR-656-3p组为模型细胞分别转染anti-NC、anti-miR-656-3p,si-NC组、si-ATP10A组为模型细胞分别转染si-NC、si-ATP10A。
1.3MTT实验 收集处于对数期的SH-SY5Y细胞,调整细胞密度,吸取200 μl细胞悬浮液接种至96孔板中,每孔含有1×105个SH-SY5Y细胞,将Aβ25~35浓度调整至0、5、10、20 μmol/L,分别加入各组细胞中,每孔设置3个复孔,加入含10%胎牛血清的培养基培养48 h,每孔细胞中加入20 μl无血清培养基稀释的MTT溶液(5 mg/ml),37℃孵育4 h,小心弃上清,每孔细胞中加入150 μl二甲基亚砜(DMSO)孵育10 min,置于酶标仪中检测每孔细胞在490 nm处吸光值(OD490 nm值)。
1.4荧光定量聚合酶链反应(qPCR) 将细胞密度调整为1.0×103/ml接种至6孔板中,待细胞密度达60%~80%,弃去培养基,每孔细胞中加入500 μl Trizol溶液,混匀,37℃裂解10 min,提取细胞中总RNA,根据逆转录试剂盒说明书合成cDNA,以cDNA为模板,β-actin为对照,根据qPCR试剂盒说明书进行操作,反应条件设置为94℃ 3 min,94℃ 30 s,57℃ 30 s,72℃ 30 s,35个循环。实验重复3次,采用2-ΔΔCt法进行分析目的基因的相对表达量。
1.5细胞的转染 6孔板中每孔接种5×105个SH-SY5Y细胞,各组SH-SY5Y细胞转染前,更换为无血清DMEM培养基,吸取5 μl阴性对照及转染质粒分别与100 μl无血清DMEM培养基混匀,制备质粒稀释液;5 μl Lipofectamine2000与100 μl无血清DMEM培养基混匀,制备Lipofectamine2000稀释液;将质粒稀释液和Lipofectamine2000稀释液混合混匀,37℃静置20 min,形成复合物;吸取200 μl复合物加入6孔板中各细胞中,混匀,37℃培养4~6 h,更换为含10%胎牛血清的DMEM培养基继续培养。
1.6流式细胞术实验 细胞浓度调整为1×105/ml接种至6孔板中,每组设置3个复孔,吸取500 μl磷脂酰结合蛋白(Annexin)Ⅴ缓冲液加入每组细胞,每组细胞中加入5 μl Annexin Ⅴ-异硫氰酸荧光素(FITC)和5 μl碘化丙啶(PI),混匀,暗室中孵育30 min,置于流式细胞仪中检测各组SH-SY5Y细胞的凋亡率。
1.7双荧光素酶报告基因 通过TargetScan数据库预测与miR-656-3p可能结合的靶基因,发现ATP10A作为本实验的候选研究对象。将购买的ATP10A-wt和ATP10A-mut荧光素酶报告载体分别与过表达或敲低miR-656-3p共转染入细胞,收集细胞,在多功能酶标仪中检测每孔海肾荧光素酶的信号及萤火虫荧光素酶信号强度,以萤火虫荧光素酶活性作为参照,统计比值,每孔设置3个复孔,实验重复3次。
1.8蛋白质印迹(Western印迹)实验 收集1×107个SH-SY5Y细胞,加入500 μl放射免疫沉淀(RIPA)蛋白裂解液,冰上裂解30 min,提取细胞中总蛋白。吸取200 μl二喹啉甲酸(BCA)工作液测定目的蛋白浓度,加入适量4×蛋白上样缓冲液,100℃加热5 min促进蛋白质变性;根据目的蛋白分子量大小,配制不同浓度聚丙烯酰胺凝胶,置于电泳槽中,加入电泳液,吸取40 μg蛋白样品分别加入上样孔,恒压80 V,电泳至样品进入分离胶,将电压调整至120 V,电泳至目的蛋白迁移至凝胶2/3处,停止电泳;取出携带目的蛋白的凝胶,割下目的条带,蒸馏水进行清洗,剪取相同大小的聚偏氟乙烯(PVDF)膜,进行转膜,含5%脱脂奶粉的封闭液中封闭2 h;加入一抗稀释液,4℃孵育过夜;含吐温20三羟甲基氨基甲烷缓冲液(TBST)洗涤3次×10 min,加入辣根过氧化物酶标记二抗,室温孵育2 h,TBST洗涤3次×10 min,加入电化学发光(ECL)液孵育,显影、拍照,Image J软件分析目的蛋白相对灰度,重复3次。
1.9统计学分析 采用SPSS22.0软件进行t检验,单因素方差分析及SNK-q检验、方差分析。
2 结 果
2.1miR-656-3p在AD模型细胞中的表达量 不同浓度Aβ25~35显著抑制SH-SY5Y细胞的增殖(P<0.05),具有一定的浓度依赖性,在10 μmol/L Aβ25~35作用下SH-SY5Y细胞的增殖活性变化幅度较大;与0 μmol/L组相比,miR-656-3p在不同浓度Aβ25~35作用的SH-SY5Y细胞中的表达量显著降低(P<0.05),其中在10 μmol/L Aβ25~35作用下SH-SY5Y细胞中变化幅度较大;miR-656-3p的表达量随着10 μmol/L Aβ25~35作用时间的增加显著降低,在24 h变化幅度较大。见表1。

表1 不同浓度Aβ25~35及10 μmol/L Aβ25~35作用不同时间对SH-SY5Y细胞增殖活性及miR-656-3p表达量的影响
2.2过表达miR-656-3p对AD模型细胞增殖、凋亡的影响 与对照组(1.000±0.069)相比,模型组、NC组、miR-656-3p组细胞中miR-656-3p表达量显著降低(0.262±0.035、0.271±0.039、0.859±0.078,均P<0.05);与模型组相比,NC组miR-656-3p的表达量无显著影响;与NC组相比,在细胞模型中转染miR-656-3p模拟物可显著增加miR-656-3p的表达量(P<0.05);模型组细胞较对照组增殖活性显著降低、凋亡率显著增加(P<0.05),模型组和NC组细胞的增殖、凋亡无显著差异;但过表达miR-656-3p较NC组显著增加细胞的增殖活性(F组间=464.163,P组间=0.000;F时间=3 831.94,P时间=0.000;F组间×时间=157.416,P组间×时间=0.000)、降低细胞的凋亡率(P<0.05)。见图1、表2。
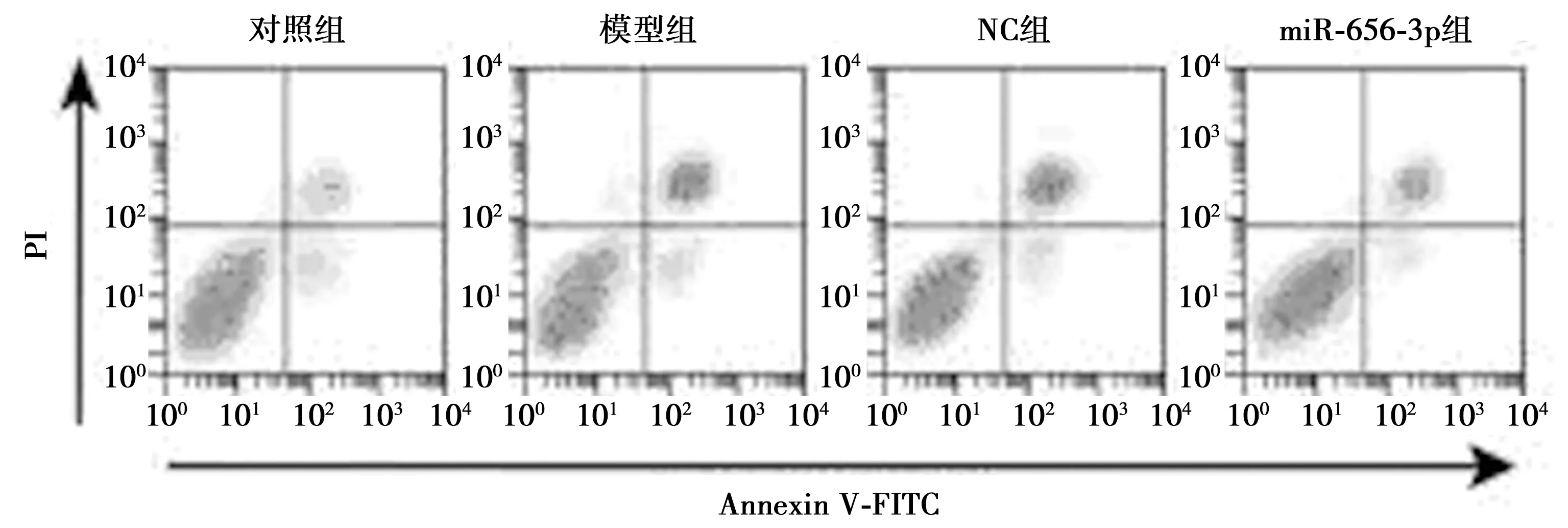
图1 各组细胞凋亡流式细胞检测
表2 转染miR-656-3p mimics对细胞增殖凋亡的影响
2.3miR-656-3p靶基因的预测和验证 在线数据库TargetScan预测miR-656-3p与ATP10A 3′UTR部分碱基可互补结合,提示ATP10A可能是miR-656-3p的下游靶基因;进一步通过双荧光素酶报告基因验证显示,与共转染ATP10A-wt和NC/anti-NC载体相比,共转染ATP10A-wt和过表达miR-656-3p/anti-miR-656-3p载体显著降低或提高SH-SY5Y细胞的双荧光素酶活性(P<0.05);与共转染ATP10A-mut和NC/anti-NC载体相比,共转染ATP10A-mut和过表达miR-656-3p/anti-miR-656-3p载体对SH-SY5Y细胞中的双荧光素酶活性均无明显的影响。与NC组相比,过表达miR-656-3p显著降低ATP10A在AD细胞模型中的蛋白水平(P<0.05);相反,下调miR-656-3p显著增加ATP10A蛋白水平(P<0.05)。见表3和图2。
表3 双荧光素酶报告基因验证miR-656-3p与ATP10A的靶向关系及蛋白表达
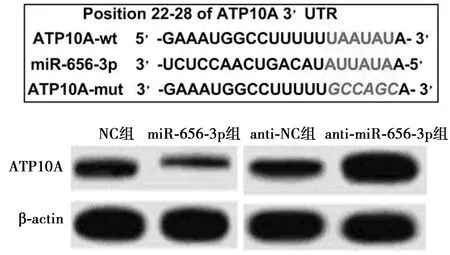
图2 miR-656-3p下游靶基因的预测及过表达或下调miR-656-3p对ATP10A蛋白水平的影响
2.4下调ATP10A对AD模型细胞增殖、凋亡的影响 与0 μmol/L组相比,ATP10A蛋白在不同浓度Aβ25~35作用的SH-SY5Y细胞中的表达量显著增加(0、5、10、20 μmol/L分别为0.126±0.009、0.228±0.013、0.569±0.042、0.731±0.051,P<0.05);ATP10A蛋白水平随着10 μmol/L Aβ25~35 作用时间的增加显著增加(0、12、24、48 h分别为0.119±0.012、0.259±0.018、0.516±0.037、0.706±0.055,P<0.05)。见图3。与si-NC组相比,下调ATP10A显著促进细胞的增殖(F组间=1 622.468,P组间=0.000;F时间=10 751.252,P时间=0.000;F组间×时间=1 315.756,P组间×时间=0.000),抑制其凋亡(P<0.05),与过表达miR-656-3p具有相似的作用。见表4。
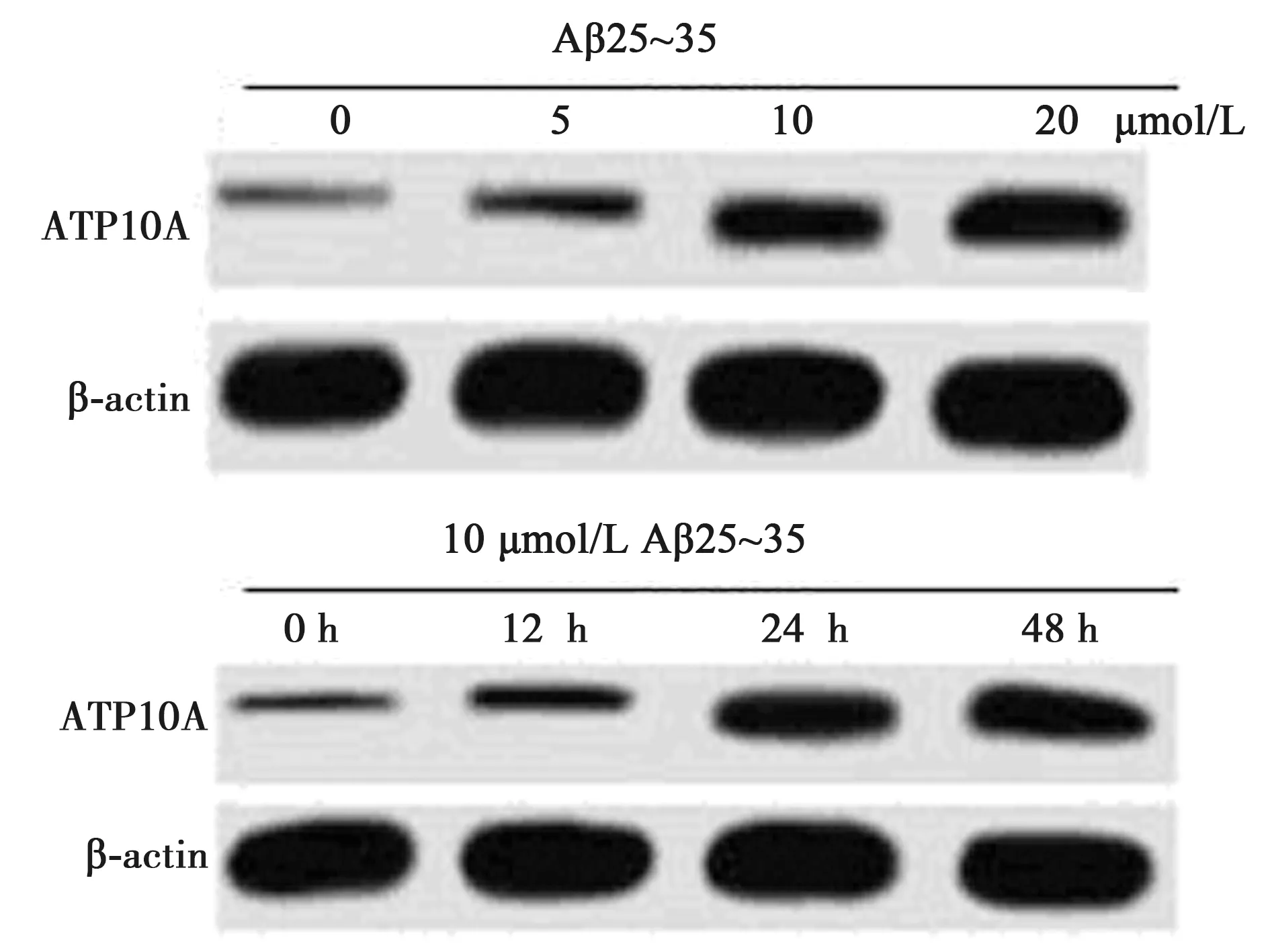
图3 下调ATP10A对AD模型细胞增殖、凋亡的影响
表4 下调ATP10A对AD模型细胞增殖、凋亡的影响
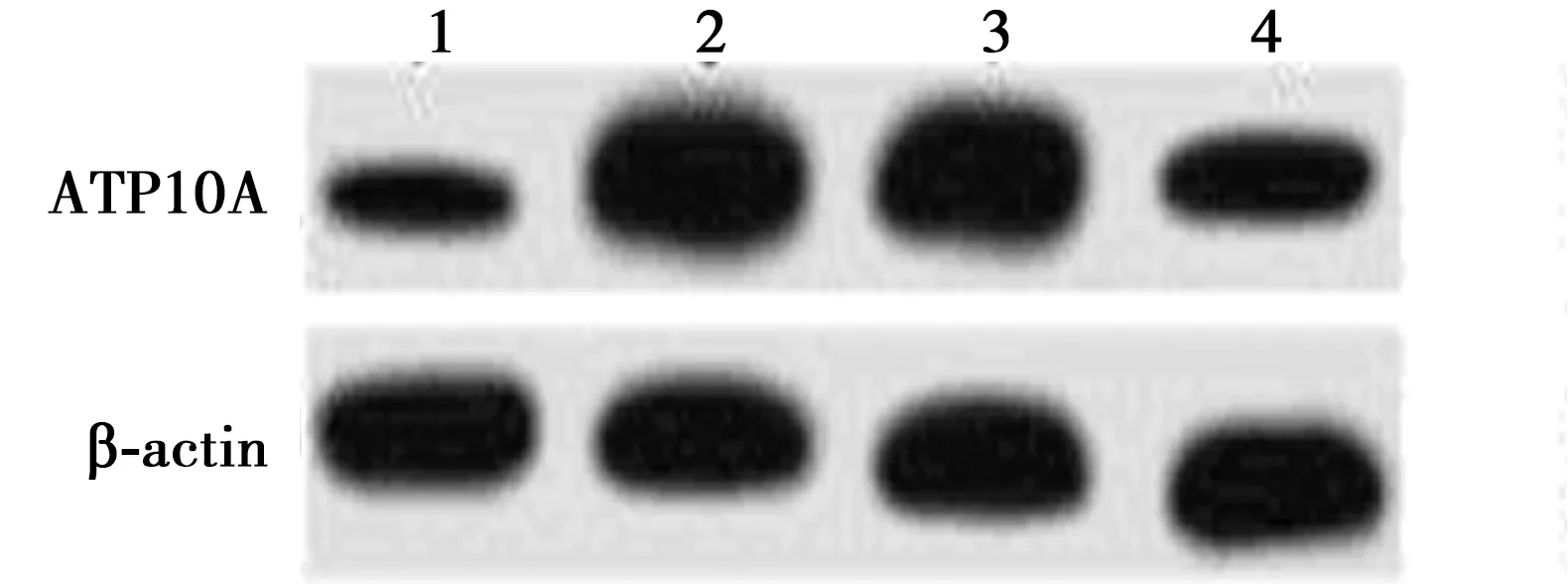
2.5过表达miR-656-3p和ATP10A对AD模型细胞增殖、凋亡的影响 与NC组相比,miR-656-3p组及miR-656-3p+pcDNA组均显著降低ATP10A的蛋白表达量,显著促进细胞的增殖,显著抑制细胞凋亡(P<0.05);miR-656-3p+ATP10A组较miR-656-3p+pcDNA组显著增加ATP10A蛋白的表达量,抑制细胞增殖(F组间=1 014.546,P组间=0.000;F时间=11 366.086,P时间=0.000;F时间×时间=288.583,P组间×时间=0.000),促进细胞凋亡(P<0.05)。见图4和表5。

1~4:NC组、miR-656-3p组、miR-656-3p+pcDNA组、miR-656-3p+ATP10A组图4 过表达miR-656-3p和ATP10A对AD模型细胞增殖、凋亡的影响
表5 过表达miR-656-3p和ATP10A对AD模型细胞增殖、凋亡的影响
2.6下调miR-656-3p和ATP10A对AD模型细胞增殖、凋亡的影响 与anti-NC组相比,anti-miR-656-3p组及anti-miR-656-3p+si-NC组均增加ATP10A的蛋白表达量,抑制细胞的增殖,促进细胞凋亡,差异具有显著性(P<0.05);anti-miR-656-3p+si-ATP10A组较anti-miR-656-3p+si-NC组显著降低ATP10A蛋白的表达量,促进细胞的增殖(F组间=1 518.042,P组间=0.000;F时间=2 168.350,P时间=0.000;F组间×时间=152.894,P组间×时间=0.000),抑制细胞的凋亡,差异具有统计学意义(P<0.05)。见表6和图6。
表6 下调miR-656-3p和ATP10A对阿尔茨海默病模型细胞增殖、凋亡的影响
1~4:anti-NC、anti-miR-656-3p、anti-miR-656-3p+si-NC、anti-miR-656-3p+si-ATP10A图6 下调miR-656-3p和ATP10A对阿尔茨海默病模型细胞增殖、凋亡的影响
3 讨 论
miR在人类基因表达量的调控具有举足轻重的作用,近年来的研究结果表明其表达量与多种疾病其中包括神经退行性疾病的演进密切相关。多种miR在心血管〔10,11〕、糖尿病〔12,13〕、肿瘤〔14,15〕、AD〔16〕等的表达量显著异常。Bekris等〔17〕研究发现AD患者脑组织、脑脊液及血液中miR-15a表达量下调。miR-34a在APPswe/PSΔE9模型小鼠中表达量显著增加,可能通过调控Bcl-2参与AD发生〔18〕。miR-124-3p可以通过靶向下调Caveolin-1表达,抑制AD细胞凋亡,降低细胞中游离钙离子浓度,从而发挥神经保护作用,为AD防治提供新的思路和靶点〔19〕。miR-153可负调控靶基因下游信号分子糖原合成酶激酶(GSK)-3β的表达;过表达miR-153可降低细胞增殖活性,增加其凋亡水平〔20〕。上述研究结果均表明miR可通过调控下游靶基因mRNA的表达量影响AD的发生发展过程。其中,miR-656在神经胶质瘤进展过程发挥抑制作用,miR-656在神经胶质瘤组织及细胞系中的表达量显著降低;体内和体外实验结果显示miR-656可抑制神经胶质瘤的恶性生长,抑制动物模型中肿瘤的生长,其作用机制主要通过调控骨形成蛋白受体(BMPR)1A参与神经胶质瘤的生长、发展〔21〕。研究发现miR-656在AD中的表达量显著异常,提示miR-656可能参与AD发生发展过程〔9〕。
AD是人类神经系统疾病,研究样品来源有限,病因尚不清楚,因此构建良好的研究模型具有重要的意义。AD动物模型主要有损伤型、自然衰老型、转基因型,每一类动物模型均可在一定程度模拟AD的临床病理特征,但均具有各自的局限性。对于探究AD分子机制的相关研究,细胞模型优于动物模型。体外培养的细胞模型具有简单、实用的特点,因此被广泛应用于AD神经毒性机制研究中。实验常用的体外培养的神经细胞,经过Aβ AD进行诱导构建细胞模型最常见,是AD较为理想的细胞模型。
本实验结果提示miR-656-3p在AD中起诱导细胞增殖、抑制凋亡的作用,从而抑制AD进一步恶化;ATP10A是miR-656-3p的下游靶基因之一,且二者表达量呈负相关;ATP10A在细胞模型中表达量显著增加,与过表达miR-656-3p的作用相似,敲低ATP10A可显著促进模型细胞的增殖,抑制细胞的凋亡,在AD中发挥保护作用;共转染过表达miR-656-3p 和ATP10A显著恢复miR-656-3p对ATP10A蛋白表达量的抑制作用,逆转miR-656-3p对细胞模型增殖的促进及细胞凋亡的抑制作用;相反,共转染敲低miR-656-3p 和ATP10A显著逆转anti-miR-656-3p对ATP10A蛋白表达量的促进作用,恢复anti-miR-656-3p对细胞模型增殖的抑制及细胞凋亡的诱导作用;表明miR-656-3p调控AD模型细胞的生长、凋亡,其作用机制是直接靶向ATP10A。
