赤霉素及萌发密度对戟叶酸模种子萌发的影响*
2020-11-09朱璧如李雨桐
朱璧如 李雨桐 孙 成
(北京师范大学生命科学学院,生物多样性与生态工程教育部重点实验室 北京 100875)
有性繁殖是植物个体实现适合度最大化的重要途径[1]。利用分子标记鉴定亲代和子代基因型,进而通过亲本分析等方法确定植物个体水平上的适合度是植物交配系统和性系统进化研究中的重要内容[2]。因此,获取亲代和子代基因型数据是衡量适合度的基础。母本基因型一般可通过提取母本叶片或种皮基因组DNA 获取,但子代基因型一般不能通过直接提取种子基因组DNA 获取。
被子植物的种子可分为有胚乳和无胚乳2 种类型。对于无胚乳种子,去除由母体组织发育而来的种皮之后,可直接提取胚的基因组DNA,用于获得子代基因型数据。但是,对于有胚乳种子,由于植物胚囊发育类型的多样性,极核和卵细胞的基因型不一定相同,所以由受精极核发育而来的胚乳和由受精卵发育而来的胚的基因型不一定相同。因此,在没有确定植物的胚囊发育类型前,直接利用种子检测子代基因型时需要将胚和胚乳分离。有胚乳种子的胚一般较小,很难将其与胚乳完全分离,直接提取胚的基因组DNA 有较大的困难。种子萌发出幼苗后再提取幼苗的基因组DNA是一个可能的解决方案。然而,利用萌发的幼苗检测子代基因型可能会受到生活史早期近交衰退(early-acting inbreeding depression)的影响[3]。例如,自交产生的种子萌发率可能会低于异交产生的种子,从而影响植物个体适合度估计,例如低估自交率。因此,利用幼苗估计子代基因型需要保证种子的高萌发率。
种子的萌发率可能受到种子休眠、个体间他感作用等因素的影响。自然休眠的种子一般可通过湿润、低温处理或植物生长激素打破休眠[4]。赤霉素是多种植物生长发育过程中的重要生长调节激素[5],具有促进种子发芽[6]、提高萌发质量[7]等作用。此外,种子萌发过程中可能会释放某些化学物质,通过他感作用对周围幼苗产生影响[8]。当萌发密度较大时,邻体幼苗周围可能会积累更多的化感物质,遭受更强烈的他感作用,萌发和幼苗生长可能会受到更强烈的抑制。
戟叶酸模(Rumex hastatusD.Don)是雌全异株(gynodioecious)的多年生灌木,是研究植物性系统进化的理想材料。本文以戟叶酸模为研究材料,设置有、无赤霉素和高、中、低萌发密度双因素变量,比较不同萌发条件下戟叶酸模种子萌发率、出土子叶长度、首粒种子萌发时间和半数种子萌发时间的差异,获得戟叶酸模最优种子萌发条件。
1 材料和方法
1.1 研究材料 戟叶酸模是蓼科酸模属多年生风媒传粉灌木,植株高度一般为90~120 cm;圆锥花序,花期4—5月;瘦果卵形,具3 棱,长约2 mm,果期5—6月。在自然种群中同时存在雌性个体和两性个体,表现为雌全异株的性系统[16]。2012年在昆明采集自然种群种子,自然干燥后置于布袋中常温保存。
1.2 种子萌发 选取约10 000 粒种子,平均分成2 份。将2 份种子分别浸泡在无菌水中,置于4℃冰箱进行低温处理,每24 h 更换一次无菌水,连续处理3 d。第4 天,将其中一份使用1×10-4mol/L 的赤霉素溶液4℃低温处理,另一份继续使用灭菌水4℃低温处理;之后继续使用灭菌水对2 份种子进行4℃低温处理,每24 h 更换一次无菌水,连续处理3 d。将前述处理的种子置于垫有2 层滤纸的培养皿(直径9 cm)中,摆放密度分别为16 粒/皿(0.063 个/cm2)、32 粒/皿(0.126 个/cm2)、48 粒/皿(0.189 个/cm2)。3 种密度水平和有、无赤霉素组合成6 种处理,每种处理设置30 个重复,置于温室中进行萌发,温室白天温度为20~35℃,湿度为20%~40%。保持培养皿中滤纸湿润。每天对培养皿的摆放位置进行随机化处理,并记录萌发种子数,测量萌发5 d 后的出土子叶长度。将第15 天的萌发率作为最终萌发率用于数据分析。
1.3 数据分析 用R 软件进行统计分析。将每一个培养皿作为一个观察样本,样本大小为180。因为萌发率是比例数据,因此使用广义线性模型(generalized linear models)分析萌发密度和赤霉素处理对种子萌发率的影响,模型中解释变量为萌发密度和赤霉素,同时考虑二者的交互作用,使用R 软件中的glm()命令。用双因素方差分析(twoway ANOVA)分析萌发密度和赤霉素处理对出土子叶长度的影响,将萌发密度和赤霉素作为解释变量,同时考虑二者的交互作用。为了检验萌发密度和赤霉素处理是否会影响种子萌发的时间,将同一种处理的30 个培养皿作为一个观察样本,通过记录每天每个培养皿中种子萌发的数量,计算每天每种处理中第1 粒种子萌发的培养皿个数(首粒种子萌发皿数),以及有50%种子萌发的培养皿的个数(半数种子萌发皿数)。这2 个指标均为随时间重复测量的计数数据,且相同处理中不同培养皿的种子萌发可能存在差异,因此采用广义线性混合模型(generalized linear mixed-effect models)分析萌发密度和赤霉素对这2 个指标的影响,萌发密度和赤霉素处理作为固定因子,时间和培养皿编号作为随机因子,使用R 软件lme4 软件包中的glemr()命令。
2 实验结果
2.1 赤霉素及萌发密度对种子萌发率的影响 无赤霉素处理时,低、中、高3 种密度下戟叶酸模种子的萌发率分别为(54.17±3.85)%、(62.19±3.53)%、(55.90±2.85)%(均值±标准误,n=30);有赤霉素处理时,低、中、高3 种密度下种子的萌发率分别为(54.79±4.11)%、(59.48±2.42)%、(61.46±1.96)%(图1)。广义线性模型分析的结果表明赤霉素(t=-0.245,P=0.807)和萌发密度(t=-0.819,P=0.414)对戟叶酸模种子的萌发率均没有显著的影响,且二者之间也不存在交互作用(t=1.122,P=0.263)。

图1 不同萌发密度和赤霉素处理下戟叶酸模种子的萌发率
2.2 赤霉素及萌发密度对出土子叶长度的影响 无赤霉素处理时,低、中、高3 种密度下出土子叶的长度分别为(5.52±0.056)mm、(5.44±0.034)mm、(5.39±0.041)mm(n=30);赤霉素处理时,低、中、高3 种密度下出土子叶长度分别为(5.86±0.072)mm、(5.86±0.050)mm、(5.95±0.044)mm(图2)。双因素方差分析的结果表明萌发密度对出土子叶长度没有显著的影响(F=0.402,df=2,P=0.669),但是赤霉素对出土子叶长度有显著影响(F=109.2,df=1,P<0.001),赤霉素处理显著提高了出土子叶长度,萌发密度和赤霉素之间没有交互作用(F=2.62,df=2,P=0.076)。

图2 不同萌发密度和赤霉素处理下戟叶酸模种子的出土子叶长度
2.3 赤霉素及萌发密度对种子萌发时间的影响 萌发处理后的第3 天,即4月1日,开始有第1 粒种子萌发,各处理组的第1 粒种子萌发的时间主要集中在4月2日(图3)。广义线性混合模型分析的结果表明萌发密度(z=-0.362,P=0.717)和赤霉素处理(z=-1.025,P=0.305)对首粒种子萌发皿数均没有显著影响。说明赤霉素和萌发密度对每个培养皿中第1 粒种子萌发的时间均没有显著的影响。

图3 不同萌发密度和赤霉素处理下首粒种子萌发皿数随时间的变化
萌发处理后的第6 天,即4月3日,开始出现萌发种子数超过50%的培养皿,从3日至9日不同处理组中萌发种子数量超过50 %的培养皿数增加较快,之后增加较为平缓(图4)。广义线性混合模型分析的结果表明,萌发密度对半数种子萌发皿数没有显著影响(z=1.604,P=0.109),但赤霉素对半数种子萌发皿数有显著影响(z=-4.025,P<0.001),赤霉素处理显著推迟了每个培养皿中种子萌发数量超过50%的时间。
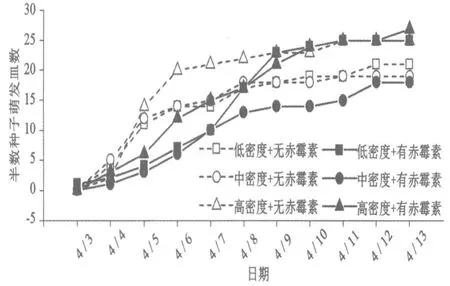
图4 不同萌发密度和赤霉素处理下半数种子萌发皿数随时间的变化
3 讨论
本实验结果表明,萌发密度对戟叶酸模种子萌发的4 个指标均没有显著的影响;赤霉素处理对萌发率和首粒种子萌发时间没有显著的影响,但经过赤霉素处理后,出土子叶长度显著增加,半数种子萌发时间显著推迟(图1~图4)。
在波动的自然环境中,植物种子可通过休眠降低所有种子同时萌发带来的潜在危险,同时植物种子还可通过捕捉特定信号改变萌发时间[9],以便在出现适宜环境的时候进行萌发,提高存活率。种子密度[10]、赤霉素[5]等都会影响种子的萌发。一般认为种子密度增加会抑制种子的萌发,因为此时种子可能通过化感物质识别到种内竞争加剧而选择休眠。Murray[9]研究了萌发密度对12 种植物种子萌发率的影响,发现11 种植物的种子在高密度(0.22 个/cm2)情况下的萌发率显著小于低密度(0.022 个/cm2)情况下的萌发率。但也有研究发现,水车前(Ottelia alismoides)种子的萌发率随着萌发密度(0.177~1.77 个/cm2)的增加而增大,二者呈显著的线性正相关关系[10]。本研究的结果显示,萌发密度的增加对戟叶酸模种子的萌发率没有显著影响(图1)。戟叶酸模耐贫瘠,耐干旱,常作为石灰岩地区的先锋植物出现[11],对生长环境和资源要求不高;此外,戟叶酸模种子较小,单个长度在2 mm 左右。因此,本研究中设置的高密度(0.189 个/cm2)对戟叶酸模种子来说可能尚未达到加剧种内竞争的程度。
种子萌发也受激素的调节,主要是受脱落酸及赤霉素的影响。脱落酸是引起种子休眠和维持种子休眠的正调节物,而赤霉素会促使合成或激活细胞壁松弛酶,诱导胚根突出从而促进种子萌发,所以赤霉素的存在可中和脱落酸的抑制作用[12-13]。但赤霉素并非对所有种子的萌发都有促进作用。有研究发现多种赤霉素浓度(0~2.29×10-3mol/L)都对巴天酸模(R.patientia)种子的萌发具有抑制作用[14]。在对小酸模(R.acetosella)种子的萌发进行研究时发现,随赤霉素的浓度从低到高变化(0~0.9×10-3mol/L),萌发率先上升后下降[15]。本研究中,赤霉素处理没有显著提高种子的萌发率,可能是因为1×10-4mol/L 的赤霉素增量并没有显著提高种子内赤霉素的浓度,因此,有、无赤霉素处理组之间的差异并不显著(图1)。
有许多研究指出,子叶的面积减小会影响小苗和周围植被之间的相互作用,同时会减少幼苗的生物量甚至降低幼苗的存活率[16]。在营养缺乏的区域,子叶的大小对小苗初期的建立意义重大[17]。赤霉素能显著提高种子萌发质量,因为赤霉素可诱导一系列与水解酶有关的基因表达,促进水解酶特别是α-淀粉酶的合成,使胚乳中的营养转化为可溶解状态供胚的萌发使用[18],促进根的伸长、增加叶面积等[5,19],从而在对阳光和水分的竞争中表现出优势[20]。本研究发现,1×10-4mol/L 的赤霉素可显著提高萌发种子的出土子叶长度(图2),不仅有利于植株的建立,更可在随后的亲本分析中提供更多的分子材料,这为植物亲子鉴定研究提供了便利。
赤霉素还可能加快种子的萌发,例如在对金刚鹦鹉棕榈籽(Macaw palm)的萌发实验中,添加赤霉素不仅让种子萌发的比例增加,而且加快了种子萌发的速度[12]。本实验中,赤霉素对种子整体的初始萌发日期没有显著影响,但显著延迟了种子整体的半数萌发时间(图3、图4),这说明赤霉素的存在减缓了种子前期萌发速率。这可能是由于萌发速度和萌发质量之间存在着权衡(tradeoff),即更好的萌发质量可能需要更长时间的萌发前准备,例如本实验中赤霉素的添加虽然减缓了种子萌发速度,但增加了出土子叶长度,提高了萌发质量。
