重新认识线粒体代谢关键酶与肿瘤的关系*
2020-11-09徐亚娟丘晓奕汪海涛薛秀花赵丽辉
徐亚娟 丘晓奕 季 鹏 汪海涛 薛秀花 杨 琼* 赵丽辉***
(1 吉林省肿瘤医院 吉林长春 130012 2 北京师范大学生命科学学院 北京 100875 3 长春理工大学生命科学技术学院 吉林长春 130022 4 中国人民解放军总医院第一附属医院 北京 100048)
1926年Warburg 发现肿瘤细胞甚至在有氧条件下也主要以糖酵解的方式产生大部分腺嘌呤核苷三磷酸(ATP),从能量代谢的角度为肿瘤治疗提供新的思路。对肿瘤细胞代谢与生长调控的关系研究,为分子靶向、抑制剂甚至医疗成像引导工具的开发,提供了理论基础[1]。由于局部血管系统不能提供足够的氧气,快速生长的肿瘤容易变得缺氧,线粒体产生的ATP 在肿瘤细胞的供能系统中所占比例较低。但线粒体在促进细胞恶性转化、增殖、逃避凋亡检测及转移扩散等方面起到关键的作用[2]。线粒体产生活性氧(ROS),参与许多生理过程的调节。正常表达量的ROS 能通过蛋白磷酸化/去磷酸化、利用相关转录因子及环鸟苷酸等相关途径参与细胞增殖、分化、凋亡等相关信息的转导。正常细胞的供能依赖葡萄糖的分解代谢,而肿瘤细胞通常依赖葡萄糖和谷氨酰胺,线粒体协调二者的平衡[3]。线粒体代谢关键酶抑制剂的开发和研究显示了靶向线粒体的潜力[4]。调节线粒体代谢相关酶的活性,以调控肿瘤细胞糖酵解和氧化磷酸化的关系,是靶向治疗的新思路。本综述讨论线粒体在肿瘤发生发展进程中的关键性作用,并从线粒体代谢关键酶的角度讨论靶向线粒体治疗肿瘤的潜力。
1 肿瘤细胞的线粒体与糖酵解
缺氧诱导因子(HIF)是一种调控多种缺氧诱导基因表达的中枢转录因子,在调节肿瘤侵袭、转移、血管生成和抗化疗抗辐射等方面发挥着重要作用[5]。线粒体在缺氧条件下不能提供足够的ATP 用于细胞存活,肿瘤细胞必须上调糖酵解途径,HIF 在这一过程中起重要作用。在肿瘤细胞中还发现,激活磷酸肌醇3-激酶(PI3K)及其下游靶向分子丝氨酸苏氨酸激酶(AKT,也称为PKB)也可刺激糖酵解,它们参与了从各种细胞表面受体传递存活信号的途径。线粒体本身也可触发缺氧诱导因子1(HIF-1)及AKT 的激活。当肿瘤细胞中线粒体呼吸下调时,三羧酸循环(TCA)底物的积累可能作为刺激糖酵解的信号。当快速生长的肿瘤将其ATP 产生转移到糖酵解时,线粒体活性降低,线粒体消耗氧气减少,ATP 产量下降。一些肿瘤氧化磷酸化机制的研究发现,线粒体ATP 合成酶(β-F1-ATPase)的催化亚单位下调,且表达水平与有氧糖酵解的速率成反比。此外,寡核苷酸在肺癌中抑制氧化磷酸化显示可引发有氧糖酵解的快速增加。这一发现表明,由于抑制线粒体能量的产生,肿瘤细胞得以糖酵解。当糖酵解被抑制时,肿瘤细胞不能充分上调线粒体氧化磷酸化,表明部分线粒体功能损伤[6]。
2 肿瘤细胞线粒体代谢的改变
代谢改变是肿瘤细胞的一个重要特征和标志。线粒体是许多细胞代谢途径中重要的参与者,包括TCA 循环、游离脂肪酸β-氧化、支链氨基酸分解代谢和类固醇生成等[7]。肿瘤细胞扩增需要大量营养物质作为核酸、脂、蛋白质合成反应的能量来源和前体。S-腺苷蛋氨酸(SAM)是合成核苷酸、蛋白质、脂质和谷胱甘肽的关键,由单碳代谢(叶酸和蛋氨酸循环结合)产生,它在肿瘤中异常表达[8]。SAM 被DNA 和组蛋白甲基转移酶分别用于DNA 和组蛋白尾的氨基酸甲基化,这可能有助于在营养缺乏时维持单碳代谢。TCA 循环几乎是连接所有单个代谢途径的最重要的中枢通路,柠檬酸可进入TCA 循环供能,或在细胞质中扩散还原为草酰乙酸和乙酰辅酶A,从而进入脂质合成。Schlaepfer I R 等[9]的小鼠实验发现,靶向阻断前列腺癌脂类分解代谢,可抑制小鼠体内前列腺癌肿瘤生长。肿瘤细胞从葡萄糖中提取的丙酮酸优先转向生产乳酸,草乙酸酯和乙酰辅酶A 通过丙酮酸或联手线粒体重组柠檬酸恶性循环改变以产生乳酸。同时柠檬酸已表现出参与对化疗、肿瘤抑制药物的肿瘤抗性增强。乳酸对TCA 循环周期中间体的贡献大于葡萄糖,乳酸是大多数组织和肿瘤中主要的循环TCA 底物。Andrea Viale 等[10]研究发现,癌基因消融而处于休眠状态的潜伏性肿瘤细胞想要存活并诱导肿瘤复发,必须依赖于氧化磷酸化产生能量才能生存。这使得肿瘤细胞对营养缺乏和环境压力更有抵抗力。
3 线粒体代谢关键酶
正常代谢的小分子成分(或对映体)积累导致信号失调,从而建立致癌的环境[11],线粒体代谢关键酶的突变在此过程起重要作用(图1)。在人肿瘤细胞中发现了核编码的三羧酸循环(TCA)中异柠檬酸脱氢酶(IDH)、琥珀酸脱氢酶(SDH)、富马酸水合酶(FH)等突变。在弥漫性胶质瘤和继发性胶质母细胞瘤中,IDH 1 突变被认为是最早可检测到的基因改变之一。AML 中的IDH 突变与TET2 甲基胞嘧啶羟化酶中的失活突变相互排斥,暗示突变IDH 可能作为表观遗传重塑的真正驱动因子[12]。此外,体内正常造血细胞IDH 1 等位基因的突变引发靶细胞谱系的异常扩增[13]。IDH 1 参与柠檬酸循环,将异柠檬酸转化为α-酮戊二酸盐 (α-KG),突变型IDH 1 则将α-KG 转化为2-羟基戊二酸(2-HG)。由于与α-酮戊二酸的结构相似性,2-HG 充当α-酮戊二酸依赖性双加氧酶的竞争性抑制剂。例如2-HG 具有抑制脯氨酰羟化酶(PHD)的作用,PHD 对HIF 的降解被抑制导致HIF-1α 的激活。2-HG 也抑制TET 酶[ten-eleven translocation,是生物体内存在的一种α-酮戊二酸(α-KG)和Fe2+依赖的双加氧酶]和组蛋白甲基化酶,导致去甲基过程受阻,肿瘤细胞CpG 岛高甲基化表型。FH 和SDH 中功能缺失突变可通过产生琥珀酸盐的积累而类似地起作用,琥珀酸盐也会拮抗这些α-酮戊二酸依赖性双加氧酶的功能。FH 的丧失和富马酸盐的积累在遗传性平滑肌瘤存在,提示线粒体代谢酶在常见肿瘤发病机制的重要性[14]。在肾癌细胞中FH 基因突变导致胞内富马酸的积累,富马酸水平升高参与包括抑制抗肿瘤因子、激活致癌HIF 信号通路等相关肿瘤发展过程。富马酸也可通过琥珀酸直接修饰半胱氨酸残基形成2-琥珀酸半胱氨酸,从而抑制乌头酸酶活性表达,引起代谢改变[15]。SDH的双等位基因丢失在家族性副神经节瘤和散发性胃肠道间质瘤中存在,会产生琥珀酸的积聚,对肿瘤发生有潜在的中心作用[16]。
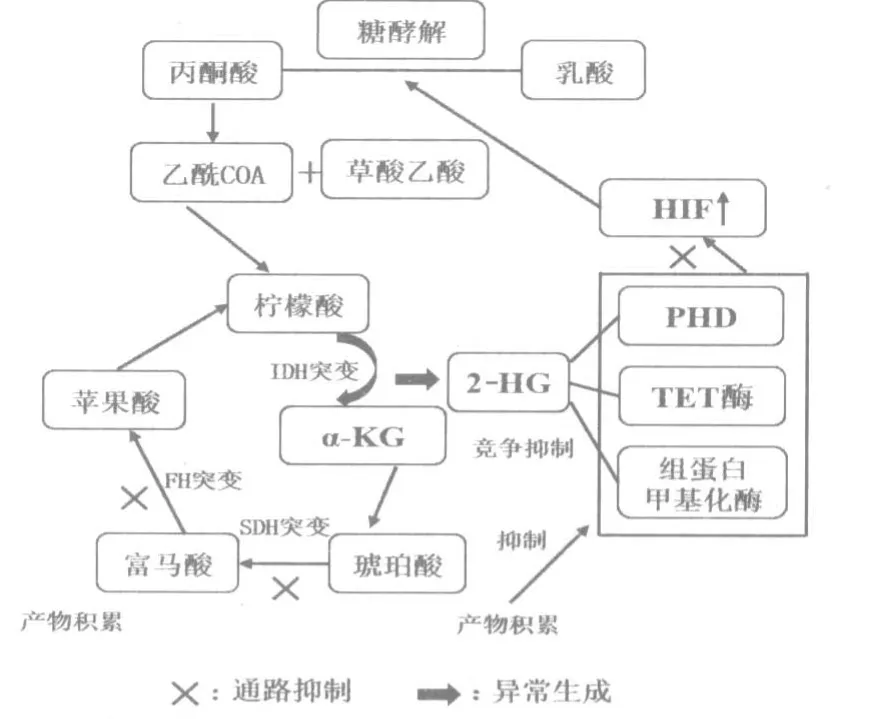
图1 线粒体代谢关键酶突变的作用途径
4 线粒体其他酶
线粒体乌头酸酶催化柠檬酸盐的立体定向异构化为异柠檬酸盐,柠檬酸介于糖酵解和脂肪酸合成的中间环节,是能量产生关键的调节器。近年来的研究阐明,胰腺癌与TCA 循环相关的顺-乌头酸和异柠檬酸的代谢改变有相关性,可作为胰腺癌患者新的预后指标[17]。α-酮戊二酸脱氢酶通过影响烟酰胺腺嘌呤二核苷酸的还原态(NADH)和ROS 的表达水平以平衡线粒体氧化还原状态,并引导TCA 代谢物通量向能量合成和信号通路的方向发展。α-酮戊二酸脱氢酶活性的变化及线粒体生物能量学的变化,不仅可能影响细胞周期通量,而且可能被放大,最终导致复杂的代谢和表观遗传重构[18]。有研究表明,在多种肿瘤细胞系中,苹果酸脱氢酶1 与谷氨酰胺的碳生成苹果酸盐,替代乳酸脱氢酶(LDH)在增殖过程中再生烟酰胺腺嘌呤二核苷酸(NAD),从而加速糖酵解的过程以维持细胞增殖所需要的能量[19]。这些参与线粒体代谢的关键酶在肿瘤细胞中的异常表达,对寻找肿瘤临床诊疗的靶点具有重大指导意义。
5 总结
自从20世纪80年代分子生物学的出现和致病性线粒体DNA 缺陷的发现以来,研究进展揭示了许多常见的人类疾病,这些疾病都与线粒体功能障碍有关。线粒体功能紊乱和线粒体内稳态紊乱在肿瘤等疾病的病理生理过程中起着重要的作用。线粒体能量代谢是细胞产能的核心途径,并满足其生物能量、生物合成和氧化还原平衡要求。尽管早期认为肿瘤细胞绕过TCA 循环并主要利用有氧糖酵解,但新的证据表明,某些肿瘤细胞严重依赖于TCA 循环产生的能量和大分子。人类多种肿瘤中显示线粒体代谢关键酶突变。随着代谢酶研究领域逐渐深入,异常TCA 循环功能在肿瘤发生中的重要性及应用小分子抑制剂干扰肿瘤治疗增强循环功能的潜力开始发展。
