如何区分趋同与平行演化?*
2020-11-09朱新宇孙利军
朱新宇 孙利军
(南通大学生命科学学院 江苏南通 226007)
1 概念的界定
趋同演化(convergent evolution)指不同谱系物种独立演化出相似的特征,即具有相似的形态或功能的结构(analogous structures),这种结构不存在于这些谱系的最近共同祖先中[1]。在支序学(Cladistics)中,这种结构被称为同型异源(homoplasy)结构。同型异源结构或特征一般出现在远缘相关的类群中,是由于演化中的趋同、同功和拟态等原因而产生的非共同性起源相似性,常作为描述系统发育重建错误起因的术语[2]。反复出现的飞行器官是典型的趋同演化的例子,例如,昆虫、鸟、翼龙和蝙蝠独立地演化出飞行能力,哺乳动物的胎盘和有袋类动物的同型异源结构;在植物中也有许多趋同演化的例子,例如,一再发生的C4光合成的发育,吸引动物取食而达到散布种子目的的新鲜水果。趋同演化的反面是趋异演化(divergent evolution)。发生趋异演化的物种具有不同的结构或特征,但这些不同的结构或特征在这些物种的最近共同祖先中具有同源器官或特征[1]。食肉植物猪笼草的捕获器结构的趋异演化是一个典型的例子。适应于本地猎物的特点,食肉猪笼草的捕获器结构发生趋异演化,最终导致属内物种的分化[3];再如,传粉者驱动的甘蓝(Brassica rapa)体型和花结构的趋异演化。通过大黄蜂(bumblebees)传粉的甘蓝具有较大的体型和更芳香的花,而食蚜蝇(hoverfly)传粉的甘蓝体型较小,并且减少了花的挥发性物质的释放[4]。
平行演化(parallel evolution)与趋同演化类似,指生活在相似的生态环境,以相同方式生存的群体,在功能和解剖结构上有许多共同特征,这些共同特征可能是同型异源结构[1],即可能已经发生了趋同或者平行演化。与趋同演化不同的是,平行演化的2 个独立物种的祖先结构或特征也是相似的,2 个物种以相同的方向演化出同型异源结构。刺鱼(stickleback)是刺鱼科(Gasterosteidae)的大约12 种鱼的统称,是平行演化的典型案例。海洋刺鱼(marine stickleback)有一排连续的32~36 个的多刺盔甲板,而淡水刺鱼(freshwater stickleback)的多刺盔甲不连续,且明显减少,只保留0~9 个多刺盔甲,淡水刺鱼的多刺盔甲的减少具有选择优势[5]。平行演化与重复演化(iterative evolution)也有相似之处。重复演化(iterative evolution),又称迭代演化,指在同一演化谱系中,灭绝物种重新复活的自然过程,重新复活的物种与灭绝物种具有相似或平行的结构或特征[6]。显然,在时间上,重复演化物种具有的与祖先物种相似或平行的结构是不连续的,被中断了。重新出现的相似形态或结构可能由于某种调控基因具有的对形态特征的超级调控能力。化石记录中有许多重复演化的例子,一个典型的例子是无羽毛的白喉秧鸡(Dryolimnas cuvieri),这个物种大约在136 000年前由于海平面上升导致灭绝,但在大约100 000年前,当海平面下降后重新出现在Aldabra 岛上,目前在这个岛上仍能发 现[6]。
趋同与平行演化可通过系统发育树上的2 种不同特征状态的变化予以区分(图1)。在平行演化过程中,同型异源特征Y 2 次演化自祖先特征X(图1a),而在趋同演化过程中,同型异源特征Y演化自不同祖先特征X 和Z(图1b)。在2 种演化类型中,同型异源特征Y 都没有与最近的共同祖先共享特征。在图1a 中的X 特征是同源的,在图1b 中的Z 特征是同源的。
在分子水平上,也存在趋同演化的案例。蛋白酶内在的化学约束导致独立的,重复的趋同演化[7]。丝氨酸(serine)和半胱氨酸(cysteine)蛋白酶使用不同的氨基酸功能基团(乙醇或者硫醇)作为亲核体。为了激活亲核体,在催化中心具有定向的酸性和碱性残基,引起一致的催化中心排列,这种在酶催化中心的化学和物理约束,在不同的酶超家族中独立演化20 次,表明在这些家族中发生了趋同演化。研究发现,趋同也发生在DNA 水平,例如,非编码DNA,顺式调控元件的分子演化速率的趋同[8]。
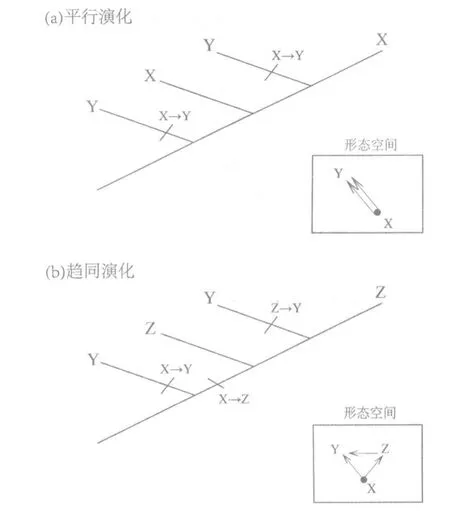
图1 平行与趋同演化的拓扑结构区分[译自Pearce(2012)[9]]
2 争议
Arendt 和Reznick(2008)[10]指出,大多数生物学家依据分类学距离区分趋同和平行演化,认为趋同是远缘分类元之间的同型异源演化,而平行是近缘分类元之间的同型异源演化。然而,这种区分没有根据,因为近缘物种常通过不同的机制演化出相同的表型,而远缘物种常通过相同的机制演化出相同的表型。大多数生物学家没有抓住趋同与平行演化的本质,即趋同主要来自外部环境的自然选择,而平行主要来自内部环境的约束。当祖先特征无法指定或未知时,在趋同和平行演化之间的区分变得更加主观。某些科学家论证,在平行和趋同演化之间存在连续性[11-12],也有些科学家认为,尽管有重叠,二者之间依然存在重要的差别[12]。
使用系统发育拓扑结构方法区分趋同和平行演化回避了导致同型异源特征的发育信息。首先,如果考虑发育信息,平行演化的祖先特征可存在不同的机理,这样依据形态的拓扑结构区分就变得不可靠;其次,不清楚2 个独立演化的物种获得的特征需要多少发育信息的重叠才能判断为同型异源特征。根据Gould 的观点[12],平行演化的概念是模糊的。从发育角度,它像趋异演化,因为支撑所产生的同型异源结构的发育机制是同源的;从选择的角度,它又像趋同演化,因为所产生同型异源结构是选择的结果。
Gould 的观点可用表1加以说明。在表1中,趋异演化形成的物种共享遗传性,它们有同源的祖先特征;平行、趋同和重复演化形成的物种是自然选择的结果,其中,趋同演化的祖先特征是非同源的,而平行演化的祖先特征是同源的。在区分趋同与平行演化时,Gould 的方法类似于同源-同型异源的拓扑结构划分,但使用了发育机制方法以区分趋同和平行演化。Arendt 和Reznick(2008)[10]批评从发育机制对趋同和平行进行解释,认为由于同型异源特征的产生涉及基因网络而不是简单的线性通路,不可能知道需要多少同源性才能判断一个同型异源案例的祖先特征是同源的或是非同源的,建议应该将所有独立演化出的同型异源结构使用趋同演化解释。然而,从认知角度,使用发育机制区分趋同和平行演化或许更为深刻。

表1 趋异、趋同、平行和重复演化比较
3 结语
区分趋同与平行演化应把握二者的本质区别,即趋同主要来自外部环境的自然选择,而平行主要来自内部环境的约束[13-14],持前者观点者被称为外部主义者(externalist),持后者观点者被称为内部主义者(internalist)。依据拓扑结构区分趋同和平行演化存在缺陷,即将平行演化产生的同型异源特征直接来源于相同的祖先特征,而忽略可能起源于不同发育机制,由此造成趋同与平行演化之间的灰色区域。由于外部-内部的争论是永久的,而目前对趋同与平行演化作出明确区分又非常困难[9],在这个意义上,拓扑学区分至少在工作层面上是有益的。笔者认为,在具体的教学和科研实践中,不必刻意在二者之间作出明确区分,正如Pearce[9]所表述的:生物学家可将趋同和平行演化应用于同一个案例,也可拒绝应用于同一个案例。
