摘叶对‘赤霞珠’葡萄光合能力的影响
2020-10-29管雪强王俊芳王恒振杨东岳尚明华刘洪勇孙玉霞王世平
管雪强,王俊芳,王恒振,杨东岳,尚明华,刘洪勇,孙玉霞,王世平*
(1. 山东省农业科学院农产品研究所/山东省农产品精深加工技术重点实验室/农业部新食品资源加工重点实验室,山东济南 250100;2. 山东省农业科学院科技信息研究所,山东济南 250100;3. 中粮长城葡萄酒(蓬莱)有限公司,山东烟台 265608)
摘除果实区域叶片可以改善果实微域环境的光照条件,从而提高果实香气、单宁、花色苷等风味物质含量[1,2]。目前,摘叶已经成为酿酒葡萄生产上的常规技术,在各主要葡萄酒产区广泛应用,而摘叶的标准、方式和时间也因自然条件、品种特点的差异而有所不同[2]。目前,大多数产区一般采用转色期摘叶的方式[3],即在果实转色时摘叶以促进花色苷和香气物质的积累。由于单宁的主要合成时期是在幼果期[4],转色期摘叶对果实单宁的合成没有明显影响,因此许多产区开始将摘叶时期提早到幼果期甚至花期[5]。在早期摘叶时,由于所摘叶片建造完成时间不长,还处于功能鼎盛期,同时摘除的叶片占叶片总量的比例也比后期明显要高,因此带来早期摘叶可能极大地影响植株整体光合能力的风险。本研究的目的就是测定摘叶对葡萄叶片和枝条光合能力的影响,以期为烟台产区及类似温带大陆性季风气候的产区,构建适宜的酿酒葡萄品质调控技术。
1 材料与方法
1.1 试验材料
本试验于2016—2018年在中粮长城葡萄酒(蓬莱)有限公司基地进行,砧穗组合为‘赤霞珠/5BB’,定植时间为2012年,行株距为2.5 m×0.8 m。
1.2 试验处理
分别于花期、幼果期和转色期对果实区域进行全摘叶(摘除上部果穗以上第一节和所有以下叶片及副梢)和半摘叶(摘除下部果穗节及以下各节的叶片和副梢)处理,以不摘叶为对照。每处理设一个杆空(8株),各处理采用随机区组设计,设5个重复。
1.3 测定方法
1.3.2 单叶光合能力测定
于摘叶后当天、摘叶2 d后和摘叶20 d后,用PP systems公司产Ciras-3光合仪测定各处理叶片的单叶光合速率;光合仪叶室为方形,规格为25 mm×18 mm。测定单叶光合时,设定光强为800 μmol/(m2·s),用叶室直接夹住叶片,使叶片充满叶室进行测定。为确保使用同一个对照,在测定前2 d对部分植株的果实区域叶片先期摘除,然后在2 d后对实际处理部分的植株果实区域叶片进行摘除。采用随机区组的顺序进行光合速率的测定,以消除测定时间段不同处理所形成的系统误差。
1.3.3 枝条光合能力的测定
2016和2017年,于摘叶后当天、摘叶2 d后和摘叶20 d后,用Ciras-3光合仪测定各处理枝条的光合速率。测定时将枝条放入高100 cm、直径20 cm的圆柱形同化室中,并根据光合同化室大小,将所测得的A值换算成整个枝条的瞬时光合速率,单位为μmol/(m2·s),换算公式为:Pn=A×Area×0.014/3。
2 结果与分析
2.1 不同时期摘叶后的枝条剩余成叶数量
由表1可以看出,花前全摘叶后,成叶数量明显减少,2016、2017、2018年每个枝条的剩余成叶数量分别为6.53、4.73、8.07片。其中2017年由于春季干旱,成叶建成较少,花前全摘叶处理后,枝条剩余成叶数量最少,仅有对照的50.35%;而2018年由于春季雨水较多,枝条剩余成叶达到对照的59.02%。半摘叶处理后,3年的枝条剩余成叶数量分别为8.67、6.67、10.20片。幼果期摘叶后,枝条剩余成叶量分别为10.73、7.47、12.13片,即便是春季较干旱的2017年,剩余枝条成叶数量也达到对照的60.87%。转色期时,由于已进入雨季,主梢和副梢生长量都很大,且都已经过数次新梢修剪,果实区域全摘叶后,仅主梢剩余成叶数量就分别达15.13、15.93、16.60片。
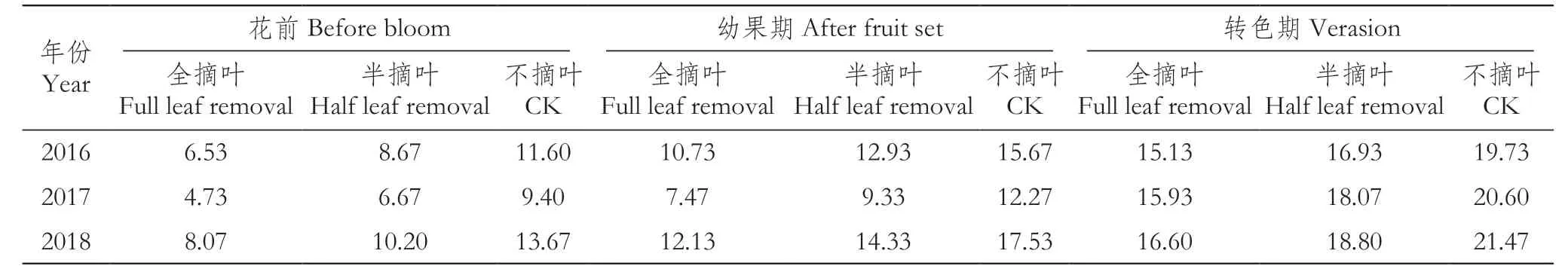
表1 不同时期摘叶后的枝条剩余成叶数量Table 1 Number of leaves left on branch after leaf removal in different periods
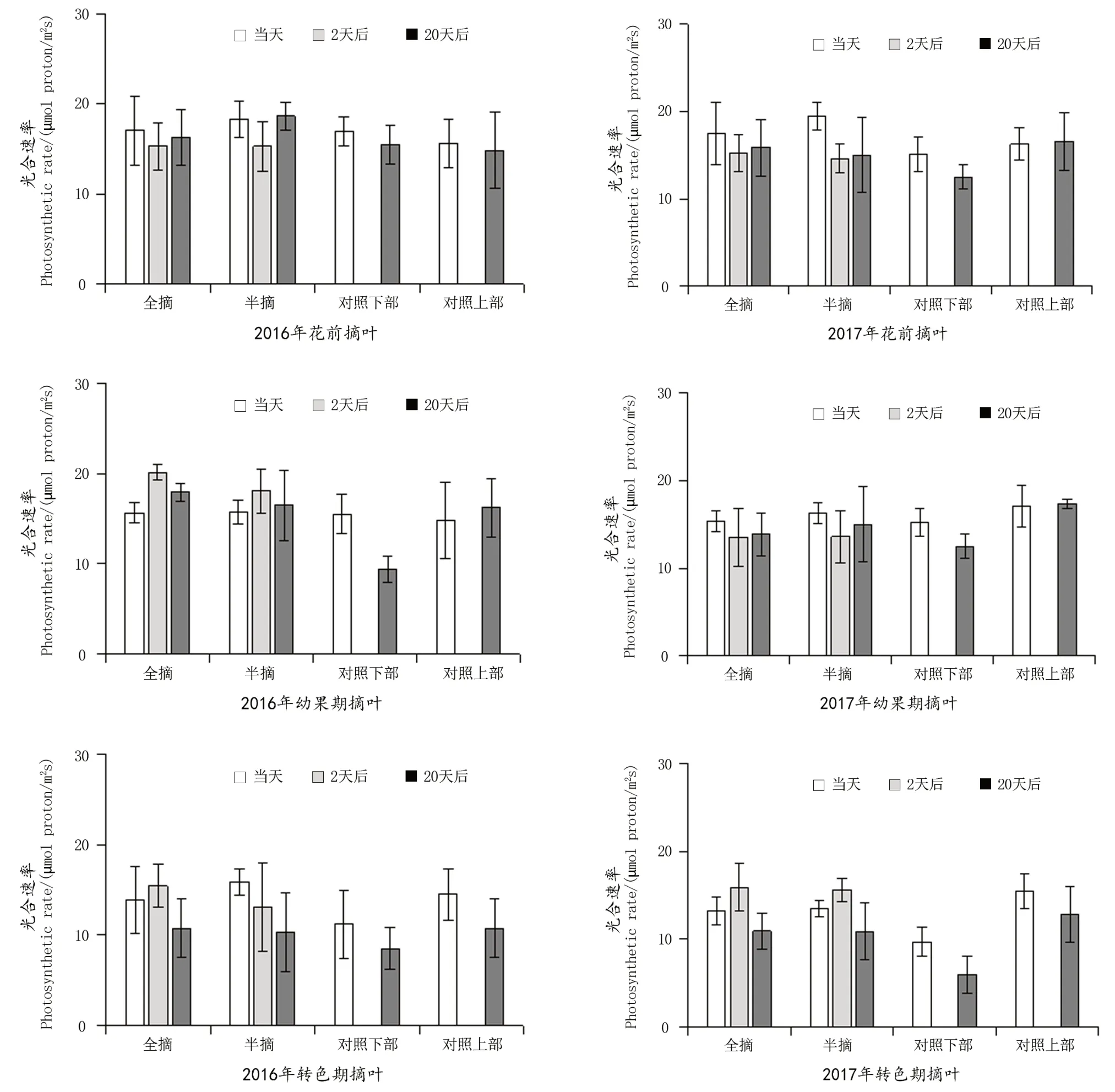
图1 摘叶对单叶光合速率的影响Figure 1 Effect of leaf removal on single leaf photosynthetic rate
2.2 不同时期摘叶对单叶光合能力的影响
由图1可以看出,2016年花前摘叶后当天剩余叶片的单叶光合速率与对照基本相同,2 d后明显低于对照,而20 d后单叶的光合速率得到恢复,甚至高于对照;2017年花前摘叶当天,剩余叶片的单叶光合速率明显高于对照,2 d后下降到与对照相当,20 d后略有恢复,且与对照无差异,分析认为摘叶2 d后叶片光合速率的下降与摘叶造成的枝条损伤有关。幼果期摘叶后,2016年和2017年剩余叶片的单叶光合速率都与对照无差异,但2016年摘叶2 d后单叶光合速率不降反升,说明摘叶对枝条形成的伤口已经不会影响到叶片功能,摘叶20 d后单叶光合速率虽有所下降,但仍高于摘叶当天的水平,与对照叶片的变化趋势基本相同;2017年幼果期摘叶后的叶片单叶光合速率变化趋势与2016年的花前摘叶基本一致,这与2017年生长前期干旱,枝条成叶数量较少有关。转色期摘叶之后,除半摘叶处理外,单叶光合速率的变化趋势在两年中都与2016年的幼果期摘叶基本一致。
2.3 不同时期摘叶对枝条光合能力的影响
由图2看出,2016年花前摘叶处理后,枝条的光合能力与对照相比明显降低,但2 d后半摘叶处理的枝条光合能力有所恢复,全摘叶处理的枝条光合能力则进一步降低,即便是20 d后,全摘叶处理的光合能力也仅有小幅回升;2017年花前摘叶处理后,全摘叶的枝条光合能力变化趋势与2016年相近,而半摘叶处理的枝条光合能力在摘叶2 d后也出现了进一步的降低,20 d后才恢复到与对照相近的水平。幼果期摘叶后,当天枝条的光合能力都是有所降低,且2 d后大都恢复到与对照相近的水平,只有2017的全摘叶处理在2 d后的恢复程度仍然与对照有较大的差距。另外,由于进入雨季后枝条生长旺盛,摘除的叶片占枝条总叶片数的比例大为降低,且枝条已经很难从架面上分离出来以测定整个枝条光合能力,因此转色期后摘叶各处理及幼果期摘叶20 d后的枝条光合能力未进行测定。
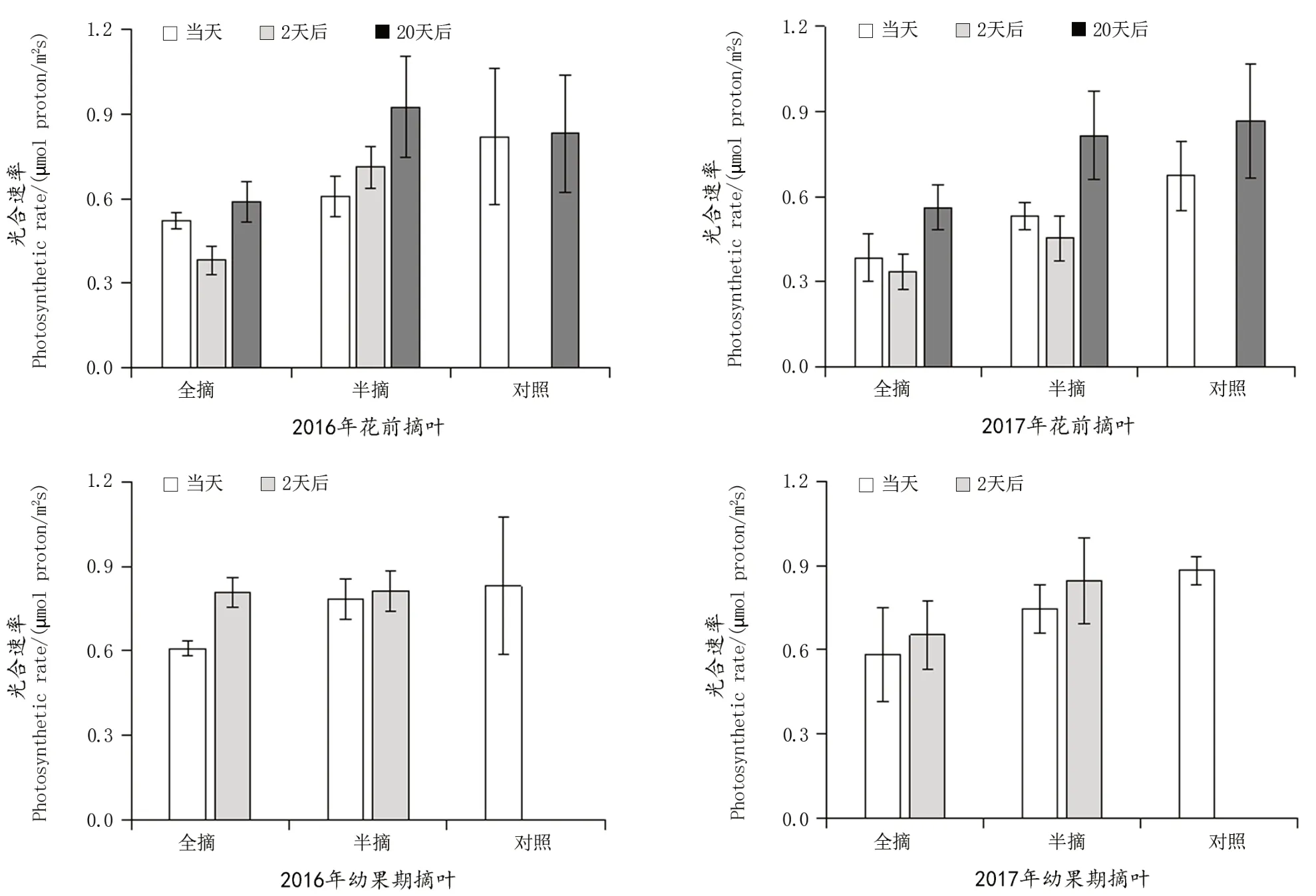
图2 摘叶对枝条整体光合能力的影响Figure 2 Effect of leaf removal on single shoot photosynthetic capacity
3 讨论与结论
摘叶能够提高果实品质这一结论已经得到了许多研究的支持[6-8],而对于光合特性的研究相对较少,且大都集中在转色期摘叶对单叶光合速率的影响上,结论也基本上都是摘叶显著提高了剩余单叶的光合速率[9-10]。但就对葡萄果实发育的影响而言,整个枝条乃至叶幕整体的光合能力比单叶光合速率更能得到直接的体现。本文结合摘叶后剩余成叶数量及单叶的光合表现,发现不同时期摘叶对单叶光合速率和枝条光合同化能力有着不同的影响,枝条发育前期摘叶(如2016年花期全摘叶和2017年的花期及幼果期全摘叶)会严重削弱单叶光合速率,并使枝条光合能力在较长时间里不能得到恢复,这不仅会影响到花期胚珠受精、种子形成及果粒的均一性,而且由于此期是黄酮醇合成的关键时期[11],还会对黄酮醇的合成造成影响,进而影响到果实品质。枝条前期建造良好的情况下在幼果期及之后摘叶(如2016年的幼果期及转色期摘叶、2017年的转色期摘叶),则会提高单叶光合速率,对枝条整体光合能力不存在不利影响。由于单宁合成的关键时期是幼果期[12],本研究认为在不严重削弱枝条光合能力的情况下尽可能早地摘叶,对单宁合成会有明显的促进作用。转色期摘叶虽然对叶片光合能力不再形成不利影响,但由于单宁合成的关键时期已经错过,也会对香气和花色苷的形成有所影响[13-14],而对单宁的影响相对较小。
综上所述认为,在胶东产区正常年份适宜的摘叶时期是幼果期,但如果因春季过度干旱造成枝条生长量不足,摘叶后枝条平均成叶数量不足8片,则应适当顺延摘叶时间。在试验进行的3年中,2016年属于春季降水正常的年份,前期营养生长量同历年平均值基本一致,2017年则属于春季极度干旱的年份,前期营养生长量严重不足,2018年则属于春季降水偏多的年份,前期营养生长量超过常年。而根据摘叶后单叶和枝条光合能力来看,2016年适宜花前半摘叶或幼果期全摘叶,2017年则只能进行幼果期半摘叶或转色期全摘叶,2018年至少可以采取与2016年相同的摘叶方式。
