甘蓝型油菜HD-ZIP基因家族的鉴定和表达分析
2020-09-10昝领兄李开祥贾永鹏杜德志
昝领兄,李开祥,贾永鹏,杜德志
(青海大学 农林科学院,青海省春油菜遗传改良重点实验室,国家油菜改良中心青海分中心,农业农村部春油菜科学观测实验站,青海省春油菜工程技术研究中心,西宁 810016)
油菜是主要的油料作物之一,由于各地环境条件有所差异,对油菜品种的特性要求越来越高,在产量高的基础上,还应具有优质、抗倒伏和抗逆性等其他特点。其中甘蓝型油菜作为主要的油菜栽培品种,因具有适应性广,产量高,品质优良等许多特性,使其种植面积最为广泛。因此揭示甘蓝型油菜抗逆性分子机理的研究对农业生产至关重要。
同源异型亮氨酸拉链蛋白(homeodomain-leucine ZIPper, HD-ZIP)是高等植物所特有的一类转录因子,其中包括C末端由61个氨基酸构成的高度保守的DNA结合结构域(Homeodomain, HD)和跟随其后的亮氨酸拉链结构域(Leucine ZIPper, LZ)[1-2]。通过LZ结构域,HD-ZIP蛋白才能折叠成为同源或异源二聚体,并且LZ结构域可以实现和其他蛋白质的相互作用,使该蛋白发挥不同的功能。此外,有研究表明含有HD-ZIP结构域的转基因植物在受到不同胁迫的环境条件下能够表现出优良的的形态特征[3-4]。根据该基因家族成员的序列相似性和结构域的不同可将HD-ZIP基因家族成员分为四大类,即HD-ZIP Ⅰ,Ⅱ,Ⅲ和Ⅳ[5-6]。这四大类都同时包含保守的HD和LZ结构域,促进了HD-ZIP蛋白的二聚化,但是这四大亚族之间在结构上也存在差异,即HD-ZIP Ⅰ和HD-ZIP Ⅱ可以识别DNA序列CAAT(A/T)ATTG,而HD-ZIP Ⅲ和HD-ZIP Ⅳ可识别的DNA序列为GTAAT(G/C)ATTAC和TAAATG(C/T)A[7-8]。此外,HD-ZIP Ⅱ还另外包含2个五组保守的氨基酸,半胱氨酸(Cys) 、脯氨酸(Pro) 、丝氨酸(Ser) 、半胱氨酸(Cy)和谷氨酸(Glu) ,被称为CPSCG域。 HD-ZIP Ⅲ和HD-ZIP Ⅳ共享一个高度结合的脂质/甾醇结合区域,称为START域[9-10]。
由于这四类亚家族成员在结构上存在差异,使得它们在各自的功能方面也有所不同。HD-ZIP Ⅰ蛋白质在植物对于多种环境条件的适应方面发挥重要作用,比如干旱,脱落酸(ABA)胁迫,盐胁迫和极端温度等。HB1属于HD-ZIP Ⅰ家族,在蒺藜苜蓿的初生根和侧根分生组织中被表达,研究发现它可参与对盐和渗透胁迫的反应[11]。在玉米HD-ZIP Ⅰ家族的Zmhdz10基因通过ABA依赖的信号传导途径调节植物的干旱和耐盐性胁迫[12]。HD-ZIP Ⅱ亚家族成员基因主要在避光或光照信号,植物发育和非生物胁迫方面发挥着重要功能[13]。例如,其家族的ATHB2、HAT2、HAT3和HAT4的表达主要受光敏色素调节,因为近红外光与远红光之间的比例会迅速诱导避光响应[14-15]。据报道,HD-ZIP Ⅲ可以作为一种调节因子参与植物细胞的横向伸长,木质部分化,生长素运输和侧器官分化[16-17]。HD-ZIP Ⅳ对植物表皮发育过程起着至关重要的作用[18];有研究发现这类蛋白质被认为参与毛状体形成,脂质代谢和角质层生物合成[19]。玉米OCL4(外细胞层4)抑制玉米形成毛状体并影响花药分裂或分化过程[20]。
综上所述,HD-ZIP蛋白在植物特异的发育方面表现出重要的作用,因此了解该家族成员的序列特征及相关生物信息学分析对于进一步挖掘并利用它们在各种植物中的功能机制必不可少。目前,HD-ZIP蛋白家族的基因功能在许多植物中都已经进行了分析和研究。如拟南芥[21]、玉米[22]、水稻[23]、杨树[24]、黄瓜[25]、大豆[26]等。但在油菜中未见相关报道。因此,本研究利用已公布的甘蓝型油菜基因组信息,通过对HD-ZIP家族基因的进行鉴定和序列分析,可为甘蓝型油菜的抗性研究及新品种培育奠定基础。
1 材料与方法
1.1 HD-ZIP基因的序列检索与鉴定
根据对HD-ZIP蛋白的相关研究,通过Pfam32.0(http://pfam.xfam.org/)数据库分别下载同源框的保守HD结构域和亮氨酸拉链LZ结构域的隐马尔可夫模型(HMM)PF00046和PF02183[27]。并使用Hmmsearch搜索工具在甘蓝型油菜基因组(http://www.genoscope.cns.fr/brassicanapus/)中寻找具有同源序列的基因[28],以在线软件NCBI-CDD(http://www.ncbi.nlm.nih.gov/Structure)对获得的蛋白序列进行结构域鉴定,选择同时含有HD和LZ结构域的蛋白作为甘蓝型油菜HD-ZIP蛋白基因家族。从甘蓝型油菜基因序列数据库中检索这些基因编码序列的注释信息,最后使用在线工具ExPASY(http://web.expasy.org/protparam)对获得序列进行氨基酸数目(aa),等电点(PI)和分子质量(u)等物理参数的统计分析。
1.2 HD-ZIP家族基因染色体定位和基因结构分析
在甘蓝型油菜基因组数据库中搜索HD-ZIP家族基因在染色体上的具体位置和每条染色体总长度,利用在线工具MapGene2Chromosome v2(http://mg2c.iask.in/mg2c_v2.0)绘制该家族基因的染色体定位图[29]。结合基因家族的gtf3文件,使用在线分析软件GSDS(http://gsds.cbi.pku.edu.cn/)绘制基因结构图,分析基因的内含子和外显子分布情况。
1.3 HD-ZIP家族基因进化树分析和motif 预测
利用Clustal[30]工具进行氨基酸序列比对。通过MEGA7.0[31]软件,以邻接法(NJ,Neighbor-Joining)构建系统发育进化树,使用poisson模式,将Bootstrap参数设置为1 000,其余参数设置为默认。并利用在线软件MEME[32](http://meme-suite.org/tools/meme)鉴定该家族基因蛋白保守基序,除了基序数量设置为20个,其余参数设置为默认。Motif分析结果使用TBtools软件下载保存[33]。
1.4 HD-ZIP家族基因之间的共线性分析
利用软件McScanX[34]对甘蓝型油菜的HD-ZIP家族基因进行共线性分析,采用Circos绘制关系图。
1.5 甘蓝型油菜HD-ZIP家族基因的表达分析
为了进一步分析和挖掘HD-ZIP转录因子在甘蓝型油菜不同组织中的表达模式,从NCBI数据库中下载已发表的转录组数据(在盐胁迫处理下,甘蓝型油菜HD-ZIP家族基因在根和叶片中的表达情况)。通过fasterq-dump软件将SRA数据转化为fastq数据,同时利用fastqc进行数据质控和过滤处理,然后利用STAR软件和甘蓝型油菜参考基因组进行比对,接着使用RSEM软件进行表达量定量分析,最后使用R软件中的pheatmap软件包绘制表达热图。
2 结果与分析
2.1 甘蓝型 HD-ZIP基因的鉴定及其染色体 定位
为了在甘蓝型油菜全基因组中鉴定所有的HD-ZIP基因家族成员和其在染色体上的分布情况,通过Pfam数据库中的PF00046 和PF02183在甘蓝型油菜基因组数据库(E-value=le-5)中对HD-ZIP家族基因进行初步的筛选,并且使用在线软件NCBI-CDD和ExPASY进行结构域鉴定,保留同时含有HD和LZ结构域的蛋白,最终在甘蓝型油菜基因组上获得了74个HD-ZIP基因成员(表1)。结果显示:74个HD-ZIP 转录因子的大小为162~691 aa,其中编码区序列和对应的氨基酸长度最短的为489 bp和162aa(BnaA08g04720D),最长的为2 076 bp和691aa(BnaC05g19660D),平均氨基酸个数为279个。蛋白分子质量(MW)为18 275.57 u(BnaA 08g04720D)~76 164.87 u(BnaC05g19660D),等电点(PI)为4.54(BnaA09g42630D)~10.06(BnaA 09g18250D)。
染色体定位结果显示:这74个基因在甘蓝型油菜19条染色体中匀有分布,此外还包括8条随机序列(图1),其中A染色体组上共有30个,除了A01上仅有1个,其余9条染色体上有2~4个不等,C染色体上共有25个,每条染色体有 2~4个,其余19个分布在8条随机染色体上。综上所述基因在染色体组上的分布不均匀。

表1 甘蓝型油菜HD-ZIP基因家族的特征Table 1 Characteristics of HD-ZIP gene family in Brassica napus
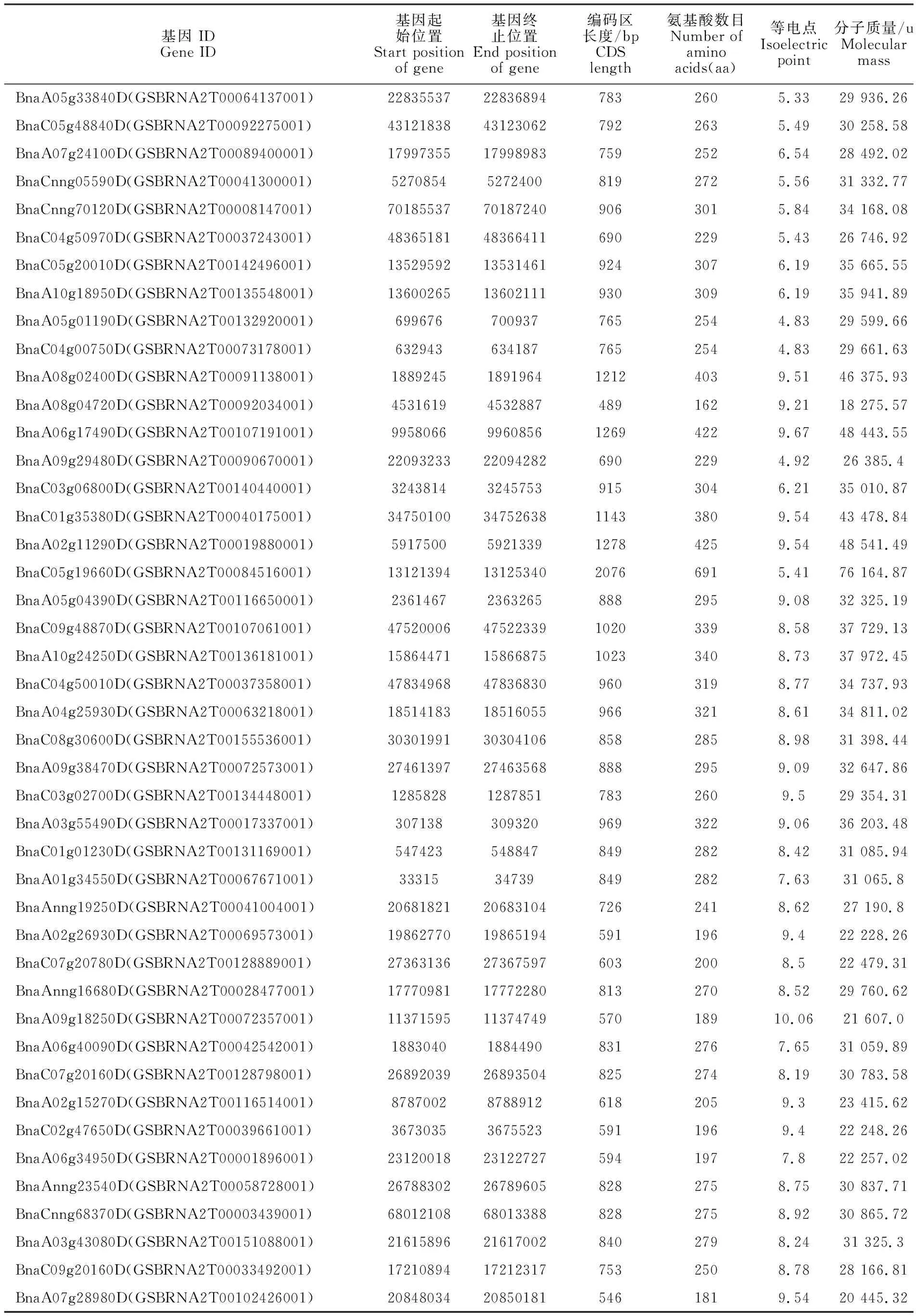
(续表1 Continued table 1)
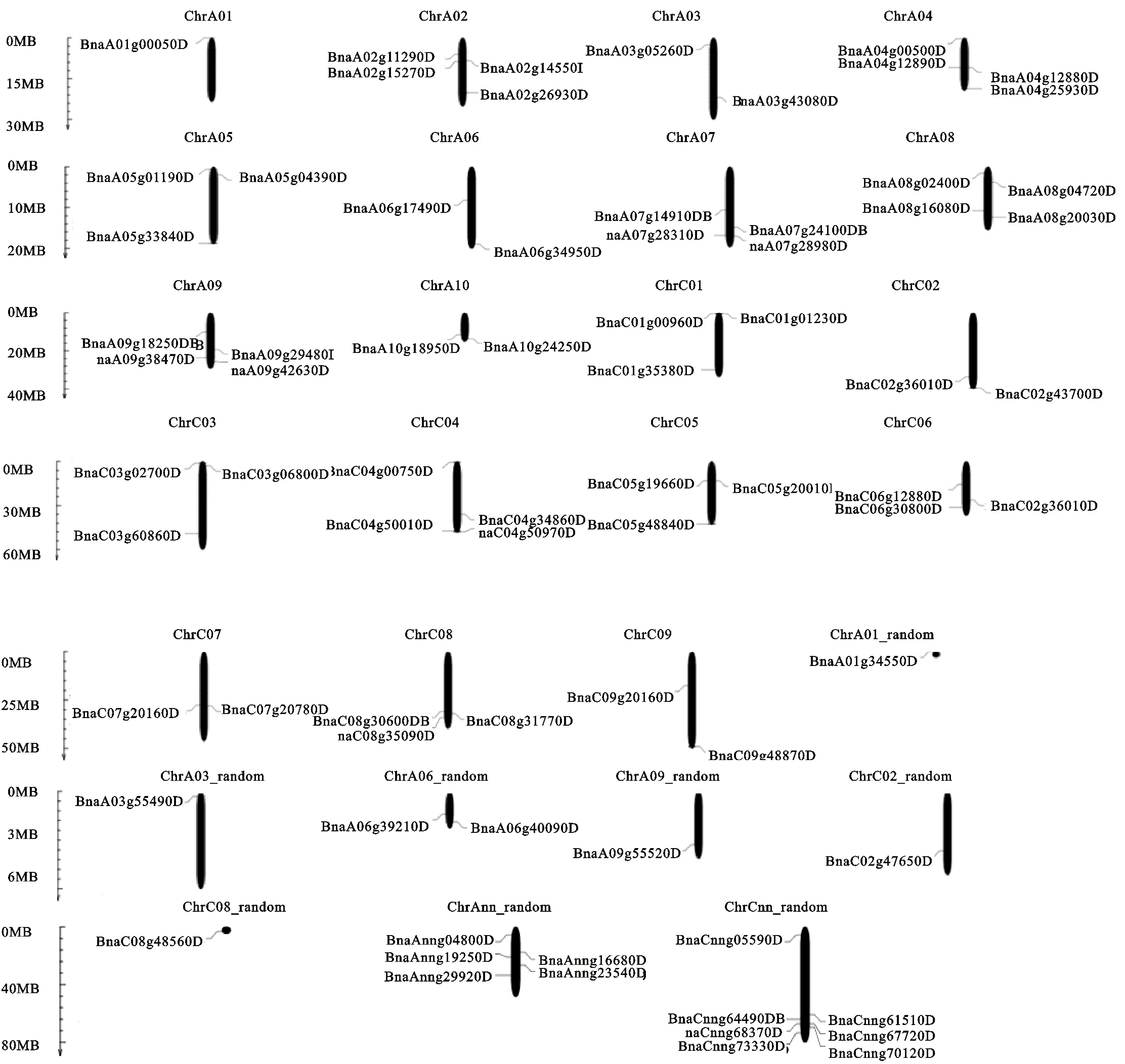
图1 HD-ZIP家族基因的染色体定位Fig.1 Mapping of HD-ZIP family gens in B.napus chromosme
2.2 甘蓝型油菜HD-ZIP家族基因系统进化树分析
为明确甘蓝型油菜中 HD-ZIP 转录因子的分类情况,本研究利用MAGA 7软件对74个甘蓝型油菜 HD-ZIP 转录因子进行系统发育进化树的构建(图2)。结果显示:74个HD-ZIP转录因子可被分为4大亚族,分别为HD-ZIP Ⅰ、HD-ZIP Ⅱ、HD-ZIP Ⅲ 和 HD-ZIP Ⅳ,其中有39个HD-ZIP转录因子被聚类在HD-ZIP Ⅰ,是最多的一类,占HD-ZIP转录因子总家族的52.7%;HD-ZIP Ⅱ和HD-ZIP Ⅳ分别占HD-ZIP转录因子总家族的9.5%和35.1%;HD-ZIP Ⅲ所包含的HD-ZIP转录因子最少,仅有2个:分别是BnaC05g19660D和BnaA09g29480D。从进化树可看出HD-ZIP Ⅰ和HD-ZIP Ⅱ处在同一分枝,其亲缘关系可能也比较相近。
2.3 甘蓝型油菜HD-ZIP家族基因结构和motif预测
使用在线软GSDS绘制HD-ZIP基因家族的基因结构图(图3),分析内含子和外显子分布情况。HD-ZIP家族基因的DNA序列和CDS分析表明:74个甘蓝型油菜HD-ZIP 的内含子个数从 1到10不等,在相同的亚族中,内含子数量差异不大,具有相似的结构。其中HD-ZIP Ⅱ的内含子数量集中为1~2,HD-ZIP Ⅳ的基因内含子个数为2~4个,基因结构比较简单;HD-ZIP Ⅰ和HD-ZIP Ⅲ的内含子个数从1~10不等;HD-ZIP家族基因的内含子在不同亚族间基因结构存在较大差异,可能是因为HD-ZIP Ⅰ和HD-ZIP Ⅲ在进化过程中内含子大量插入导致的。
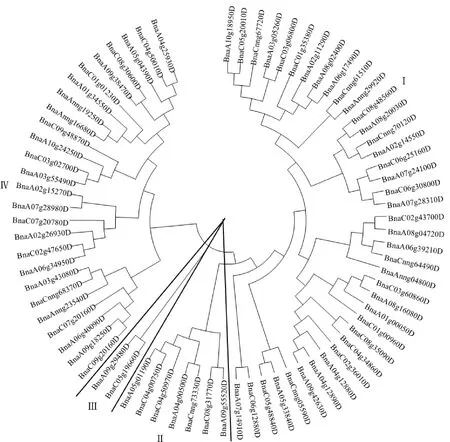
图2 甘蓝型油菜HD-ZIP转录因子的系统进化发生树Fig.2 Unrooted phylogenetic tree of HD-ZIP transcription factors in B.napus
通过MEME在线工具对甘蓝型油菜HD-ZIP转录因子的保守基序进行分析(图4)。结果显示:HD-ZIP家族的74个蛋白都含有motif 1和motif 2,这2个结构共同组成了较为保守的HD-ZIP结构域。在基因结构图和保守基序预测图中可发现同源基因间尽管在结构上存在较大的差异,但预测到的保守基序数目基本一致,比如BnaC05g19660D含有10个内含子,BnaA09g 29480D只含有3个内含子,但这2个同源基因预测到的 motif数目基本一致。但也有相反的情况,如BnaC06g12880D预测到了3个motif ,BnaC02g43700D中存在6个motif,但这2个基因都只含有2个内含子。所以同一亚族中的同源基因间基本具有相同的基序,暗示其具有的功能也可能类似。
2.4 甘蓝型油菜HD-ZIP家族共线性分析
对甘蓝型油菜的74个HD-ZIP家族基因进行串联重复性分析,挖掘HD-ZIP家族基因在甘蓝型油菜A和C基因组中的重复情况。分析结果如图5所示:鉴定到的74个HD-ZIP基因中大部分基因在甘蓝型油菜的A染色体组和C染色体组中存在重复,具有共线性关系,其中有34个基因在A和C染色体组之间具有共线性关系。同时也发现,有11个基因在A染色体组内存在共线性,5个基因在C染色体组内存在共线性。进一步说明,在异源四倍体的甘蓝型油菜基因组中,这些基因不仅在A和C染色体组组内存在同源关系,而且染色体组之间存在较多的同源性,并且存在共线性关系的基因在系统进化树中也都被聚为一类,这也表明在基因组进化过程中HD-ZIP蛋白家族基因在甘蓝型油菜中相对保守。

图3 甘蓝型油菜HD-ZIP家族基因结构Fig.3 Gene structure of HD-ZIP family in B.napus

图4 甘蓝型油菜HD-ZIP转录因子进化树和保守基序Fig.4 Phylogetic tree and gene motifs of HD-ZIP family transcription in B.napus
2.5 甘蓝型油菜HD-ZIP家族基因的表达分析
为进一步了解HD-ZIP家族基因在胁迫处理下的表达变化。结合甘蓝型油菜在盐胁迫处理下的根和叶片转录组数据。结果分析如图6所示,从图中可知,HD-ZIP家族的大部分基因在根和叶片中都发生不同程度的表达,BnaC08g35090D和BnaA09g42630D在叶片中的表达量最高,BnaA01g0050D和BnaA05g33840D在根中有最高的表达量。根中表达的基因数目多于叶片中,并且BnaA03g052600D、BnaC03g06800D等13个基因在根中的表达量要明显高于叶片,受到盐胁迫后,根和叶片中的大部分基因表达量会增加,并随着胁迫处理的时间的增加,基因的表达量又会受到一定的下调,如BnaCnng73330D和BnaA04g00500D等基因在1 h和12 h对照下表达量很低,但在盐胁迫1 h和12 h后表达量增加,并且胁迫1 h时表达量很高,但在12 h时表达量又相对较低,说明这些基因可以响应盐胁迫。只有少数基因的表达量未发生变化或者下调,如BnaC09g20160D、BnaA05g17490D、BnaA02g11290D、BnaA05g33840D。根和土壤直接接触,受胁迫后,根中的基因不断的调节变化应对外界的胁迫,然后再将胁迫的信息传递到叶片中,进行相互协调。

图5 甘蓝型油菜HD-ZIP基因的共线性分析Fig.5 Collinear analysis of HD-ZIP gene in Brassica napus genome
3 讨 论
HD-ZIP家族基因在功能上非常保守并且参与高等植物的多种生物学过程,目前大多数植物的HD-ZIP 转录因子已被鉴定和分析。Zhao等[22]在玉米中共鉴定到55个HD-ZIP家族基因,Yue等[35]在小麦中鉴定到46个HD-ZIP家族基因。但在油菜中还未见相关分析报道,为更加清楚地了解甘蓝型油菜中HD-ZIP转录因子的功能,本试验利用生物信息学方法对甘蓝型油菜基因组中的HD-ZIP家族基因进行鉴定,并对其基本理化性质、染色体定位、系统进化树,保守基序及基因结构等进行了统计分析。结果显示:在甘蓝型油菜中共鉴定到74个HD-ZIP家族的转录因子,这些基因在甘蓝型油菜19条染色体上均有分布;系统进化树将这些基因分为4个亚族;74个基因都含有motif 1和motif 2保守基序,共同组成HD-ZIP的保守结构域,这些结果与Zhao等[22]的分析结果基本一致,说明HD-ZIP家族基因在不同物种间比较保守。
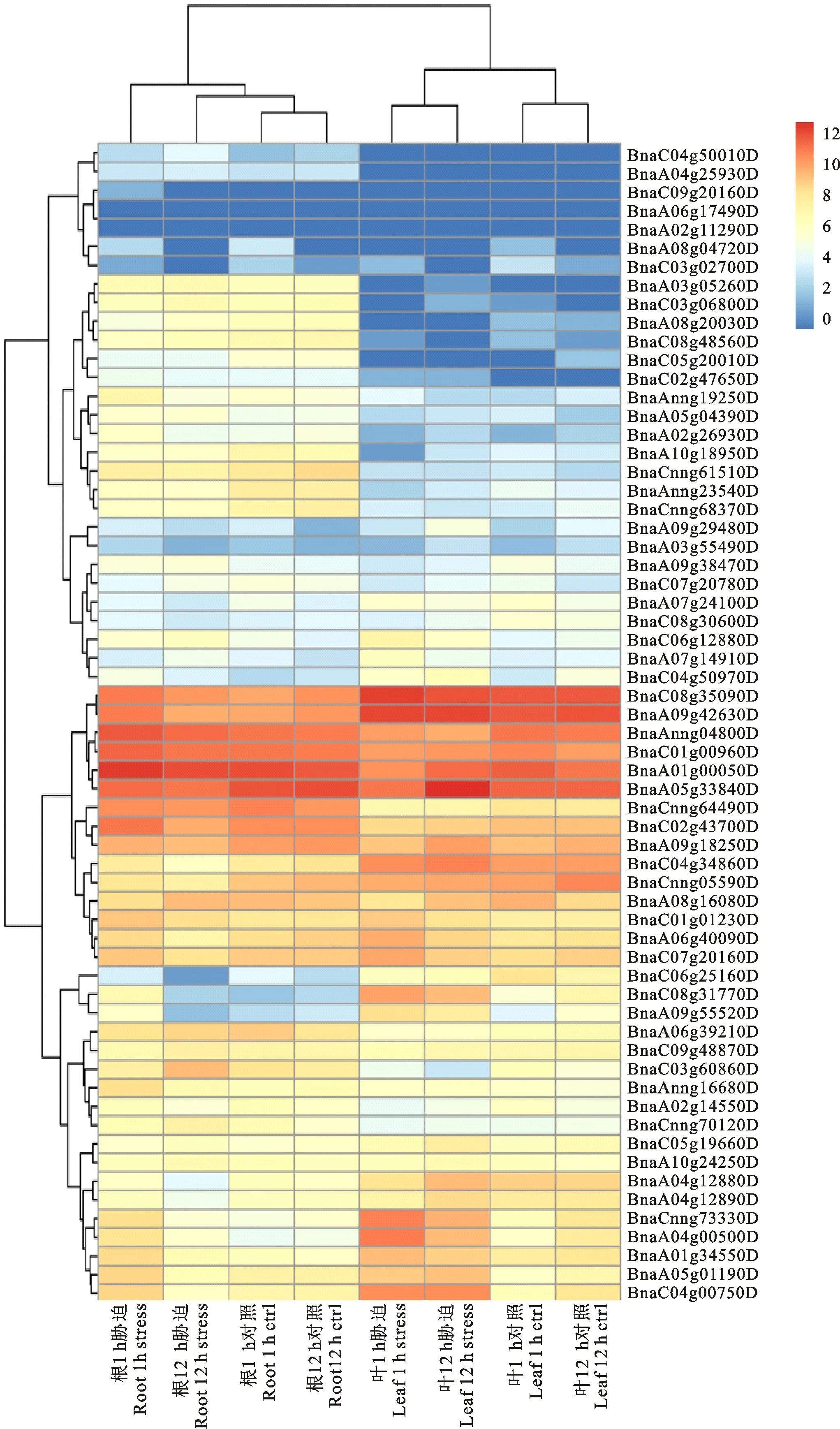
图6 甘蓝型油菜HD-ZIP家族基因的表达分析Fig.6 Expression analysis of HD-ZIP family gene in Brassica napus L.
甘蓝型油菜是由白菜和甘蓝自然杂交以及后期染色体加倍而产生的异源四倍体,基因组中应该存在多拷贝基因,但是通过共线性分析发现有的基因在甘蓝型油菜染色体组中不存在共线性关系,这可能是芸薹属植物进化时在加倍复制事件中的不对称或者丢失所引起的。但是大部分HD-ZIP家族成员基因在甘蓝型型油菜的A和C染色体组间的共线性多于在A和C染色体组内的。并且存在共线性关系的基因在系统发育进化树中也都可被聚为同一类,说明在基因组进化过程中这些基因相对保守,这可能也与这些基因在结构上存在保守的结构域有关。
HD-ZIP转录因子是一类较大的基因家族,在生物和非生物胁迫以及植物生长发育过程中具有重要的作用。在盐胁迫下对甘蓝型油菜不同组织HD-ZIP家族基因的表达模式进行分析发现,大部分基因在受胁迫时表达量会升高。Liu等[36]在拟南芥中发现BnaA04g00500D(HB12)基因对非生物胁迫(盐胁迫)响应,BnaC04g00750D(HB7)基因可以响应干旱胁迫,这些结论与本试验的分析结果一致,也进一步说明HD-ZIP家族基因在植物发育的调节和对非生物胁迫的响应中起着重要作用。同时也发现有的基因虽然为同源基因,但是在根和叶片中的表达量却相差很大,比如BnaC06g12880D、BnaA07g14910D和BnaC08g35090D、BnaA09g42630D,前2个基因在甘蓝型油菜的根和叶中表达水平低,后2个基因在根和叶中的表达量较高,但是在进化树分析中发现这两组基因都属于HD-ZIP Ⅰ亚家族,这有可能是一些基因在进化过程中功能上出现了 歧化。
