广东地区8 株鹅源基因XII 型新城疫病毒F 蛋白基因遗传进化分析
2020-08-06王杰煌朱婉君伍辉吉钟嘉诚张溢珊刘英慧张济培陈济铛
王杰煌,朱婉君,伍辉吉,钟嘉诚,张溢珊,王 贺,刘英慧,张济培,,陈济铛,*
(1. 佛山科学技术学院,广东 佛山 528225;2. 万牧洲生物科技有限公司,广东 佛山 528225)
新城疫(Newcastle disease,ND)是由ND 病毒(NDV)感染禽类引起的一种高发病率、高死亡率的急性烈性传染病[1]。NDV 分为class I 和class II 两种谱系,每一类可再分为多个基因型和基因亚型,其中,class II 至少包括18 个基因型(I-XVIII)[2]。1990 年以来,中国主要流行的是以ZJ1 株为代表株的基因VIId 亚型NDV[3-4],该亚型病毒感染家养水禽特别是鹅后呈高发病率和高死亡率特征,给我国水禽养殖业带来重大的经济损失。近年,国内禽源NDV 开始出现新的基因型和基因亚型,如2010 年,在广东活禽市场首次监测到基因XII 型NDV,随后在广东、福建等地多次分离得到该基因型病毒[5-6]。国际上,2004 年秘鲁鸡中首次发现基因XII 型NDV,随后在哥伦比亚、越南等地也陆续出现XII 型NDV感染鸡群的报道[7-8]。目前,我国XII 型分离株均属XIIb 亚型,与上述国家分离到的XIIa和XIId亚型差异较大[5-9]。本研究从广东临床呈典型NDV 感染的病死鹅中分离鉴定得到8株XII型NDV,并对这8株NDV的F基因序列进行同源性和遗传进化分析,并对其F 蛋白氨基酸序列进行分析,为进一步了解广东地区鹅源XII 型NDV 遗传进化、分子流行病学及疫苗株筛选提供参考。
1 材料与方法
1.1 病料来源临床样品采集自广东地区表现疑似新城疫临床症状的病死鹅,采集病料种类包括肝、脾、胰、脑共45份。各病毒株具体信息见表1。

表1 NDV 分离株信息表Table 1 Information of the eight NDV isolates
1.2 主要试剂DL2000 Marker、ExTaq 酶、LaTaq酶、反转录试剂盒(Prime-ScriptTMⅡ1st Strand cDNA Synthesis Kit)、 随 机 引 物(Random primer)、pMD18-T 克隆载体、大肠杆菌DH5α感受态细胞均购自TaKaRa 公司;RNA 抽提试剂盒,购自北京全式金生物技术有限公司;血凝抑制试验(HI)试验标准阳性血清均购自哈尔滨维科生物技术有限公司。
1.3 引物设计与合成参考GenBank 登录的NDV JSD0812 株F 基因序列(NC039223),利用Oligo 7 软件设计NDV RT-PCR 鉴定引物1F:5'-AGGGTCATA CATAGTCAAGT-3'/1R:5'-TAGGTGGCACGCATATTA TATC-3')及扩增病毒F 蛋白的完整编码区序列(Coding Sequence, CDS)引 物2F: 5'-TATAGTCGACAT GGGCTTCAAACCTTCTACCAG-3'/2R: 5'-TATAGCG GCCGCTCATGCTCTTGTAGTGGCTCTCA-3', 预 期 扩增片段大小分别为732 bp 和1 900 bp,引物由生工生物工程(上海)股份有限公司合成。
1.4 病毒分离鉴定将采集的病料组织研磨、冻融后,上清液经尿囊腔接种10 日龄SPF 鸡胚,收集72 h 内死亡的鸡胚尿囊液,-80 ℃保存备用。对收获的鸡胚尿囊液进行血凝及血凝抑制试验(HA/HI);利用RNA 抽提试剂盒对NDV HA、HI 效价≥4 log2 的尿囊液提取病毒RNA,反转录为cDNA 后利用1F/1R引物经RT-PCR 鉴定,PCR 产物经1%琼脂糖凝胶电泳检测。
1.5 NDV F 基因CDS 区域的PCR 扩增及测序以上述检测为NDV 阳性的cDNA 为模板,2F/2R 为引物进行PCR 扩增F 基因CDS 片段,PCR 扩增程序为95 ℃3 min;98 ℃10s,57 ℃30 s,72 ℃2 min,32个循环;72 ℃10 min。PCR 产物纯化回收后,克隆至pMD18-T 载体,构建重组质粒,以2F/2R 为引物经菌液PCR 鉴定,阳性菌液由生工生物工程(上海)股份有限公司测序。
1.6 F 基因同源性及遗传进化分析将本研究分离的NDV F 基因完整CDS 序列与GenBank 登录的NDV参考株F 基因完整CDS 序列进行比对分析其同源性,并利用MEGA 7 软件绘制该基因CDS 的遗传进化树。利用MegAlign 软件进行F 蛋白氨基酸关键位点分析。
2 结果与讨论
2.1 病毒分离和鉴定结果将病料样品上清经尿囊腔接种10 日龄SPF 鸡胚,有8 份病料组织液可致鸡胚死亡,经HA、HI 试验结果显示,8 份病料组织尿囊液HA、HI 效价均≥4 log2;RT-PCR 结果显示,扩增产物在750 bp 处出现明亮条带,与预期目的条带大小相符。上述结果表明,这8 份病料样品均感染了NDV,且经鸡胚分离到了8 株NDV(表1)。
2.2 分离病毒F 基因CDS 的PCR 扩增结果以上述检测为NDV 阳性的cDNA 为模板,2F/2R 为引物PCR 扩增F 基因CDS 片段,结果显示8 株NDV cDNA均扩增出约1 900 bp 的明亮条带,与预期目的条带基本相符。进一步表明分离得到了8 株NDV。
2.3 分离株F 基因CDS 核苷酸序列同源性分析结果分离株F 基因经测序分析,结果显示8 株NDV F 基因CDS 序列均为1 662 bp,共编码533 个氨基酸,F 蛋白裂解位点序列均为112K/RRQKR/F117,符合NDV 强毒株分子特征。F 基因核苷酸同源性分析结果显示,8 株NDV F 基因之间核苷酸同源性达96.3%~99.9%;与2010 年~2011 年广东分离到的XIIb 亚型NDV(Goose/China-GD450/2011 株)核苷酸同源性为97.4%~98.5%;与La Sota(基因II 型)、B1(基因II型)、clone 30(基因II 型)、V4(基因I 型)和Mukteswar(基因III 型)等疫苗株核苷酸同源性低,仅为83.0%~85.9%;与我国目前广泛流行的VIId 亚型NDV 代 表 株ZJ/1/00/Go 核 苷 酸 同 源 性 为88.9%~89.7%;与国内标准强毒株F48E8(基因IX 型)核苷酸同源性为85.4%~85.9%。上述结果表明,本实验分离的NDV F 基因与XII 型NDV F 基因核苷酸同源性最高,而与VIId 亚型病毒株以及常用疫苗株核苷酸同源性较低,提示广东地区鹅中出现的XIIb 亚型NDV与当前主要流行株及疫苗株F 基因差异较大。
2.4 F 基因遗传进化分析绘制本实验分离株和GenBank 下载的48 株NDV 参考株的F 基因完整CDS序列遗传进化树,结果显示,8 株新分离病毒的F 基 因 与2010 年~2011 年 广 东 基 因XII 型 鹅 源 分 离株、秘鲁、哥伦比亚以及越南基因XII 型鸡源NDV F 基因均属于class II 类。本研究分离的NDV 与2010年广东XII 型NDV F 基因同属XIIb 亚型,不同于南美病毒株(XIIa 亚型)及东南亚病毒株(XIId 亚型);与国内主要疫苗株La Sota、B1、clone 30、V4、Mukteswar4,重组新城疫病毒灭活疫苗株(A-VII 株)的亲本株NDV/ZJ1F 基因处于不同分支(图1)。结果表明,基因XIIb 亚型NDV 在广东地区鹅群中流行时间较长,正逐渐形成新的XIIb 亚型进化分支。
2.5 分离病毒F 蛋白氨基酸序列及相关位点分析比较本研究的8 株NDV 与VII 型重组灭活疫苗(A-VII株)亲本株NDV/ZJ1 的F 蛋白主要中和抗原表位氨基酸序列(aa72、aa74~aa75、aa78-aa79、aa157~aa171和aa343 位)结果显示,本研究分离的病毒株F 蛋白氨基酸序列均出现D170N 突变。本实验分离的病毒株与2011 年广东鹅中分离的XII 型病毒株(Goose/China-GD450/2011)相比,其在F 蛋白3 个疏水区(信号肽、融合诱导区、跨膜区)以及七肽重复序列[10-11]均有多个氨基酸残基出现变异,其中F399、F407 出现一致的I9L、L15P、C25H、I118V、I510T 残基替换,而F429、F438 出 现 一 致 的I10L、R18G、M20T、C25H、I118V 和I474L 变异;与V4、La Sota、Mukteswar、ZJ1和F48E8 株相比,其F 蛋白氨基酸序列差异明显,其中信号肽区域变异最明显,均出现L10P、T516I/M/G突变,与除ZJ1 株外的其余病毒相比均出现I16T、T17V、V121I 突变等(表2)。除上述区域外,本实验分离的病毒株还出现N240S、T496I 突变,该结果与2017年从广东清远鹅中分离到的Goose/CH/GD/E115/2017株的F 蛋白氨基酸变异相同[10]。上述结果表明,与常见疫苗株、主要流行株、甚至2010 年~2011 年分离到的XIIb 亚型NDV 相比,本实验分离株F 蛋白中和抗原表位等功能区域均存在差异,这些差异可能引起F 蛋白构象变化导致病毒对鹅致病力增强。值得注意的是,本实验8 株基因XII 型NDV 均分离自病死鹅,属国内首次报道。基因XII 型NDV 对鹅致病力是否出现变化,从而导致病鹅死亡有待进一步研究确定。
综上所述,目前基因XII 型NDV 在广东地区鹅群中的流行日趋广泛,部分病毒株可引起不同日龄鹅发病、死亡,严重威胁水禽养殖业。结合本研究F 基因核苷酸序列同源性分析和F 蛋白氨基酸序列分析结果,表明XII 型NDV F 基因与当前常用VIId型及其他常见NDV 疫苗株F 基因同源性较低,F 蛋白多个抗原位点氨基酸也存在差异,这些差异对基因XII 型病毒株各种生物特性,如宿主嗜性、毒力强弱、传播能力以及免疫原性等有何影响,还有待进一步研究。有必要持续关注并监测基因XII 型NDV 流行、进化情况和生物学特性变化,并进行疫苗保护效力评价,及时有针对性地调整免疫疫苗、以加强对该病的防控。
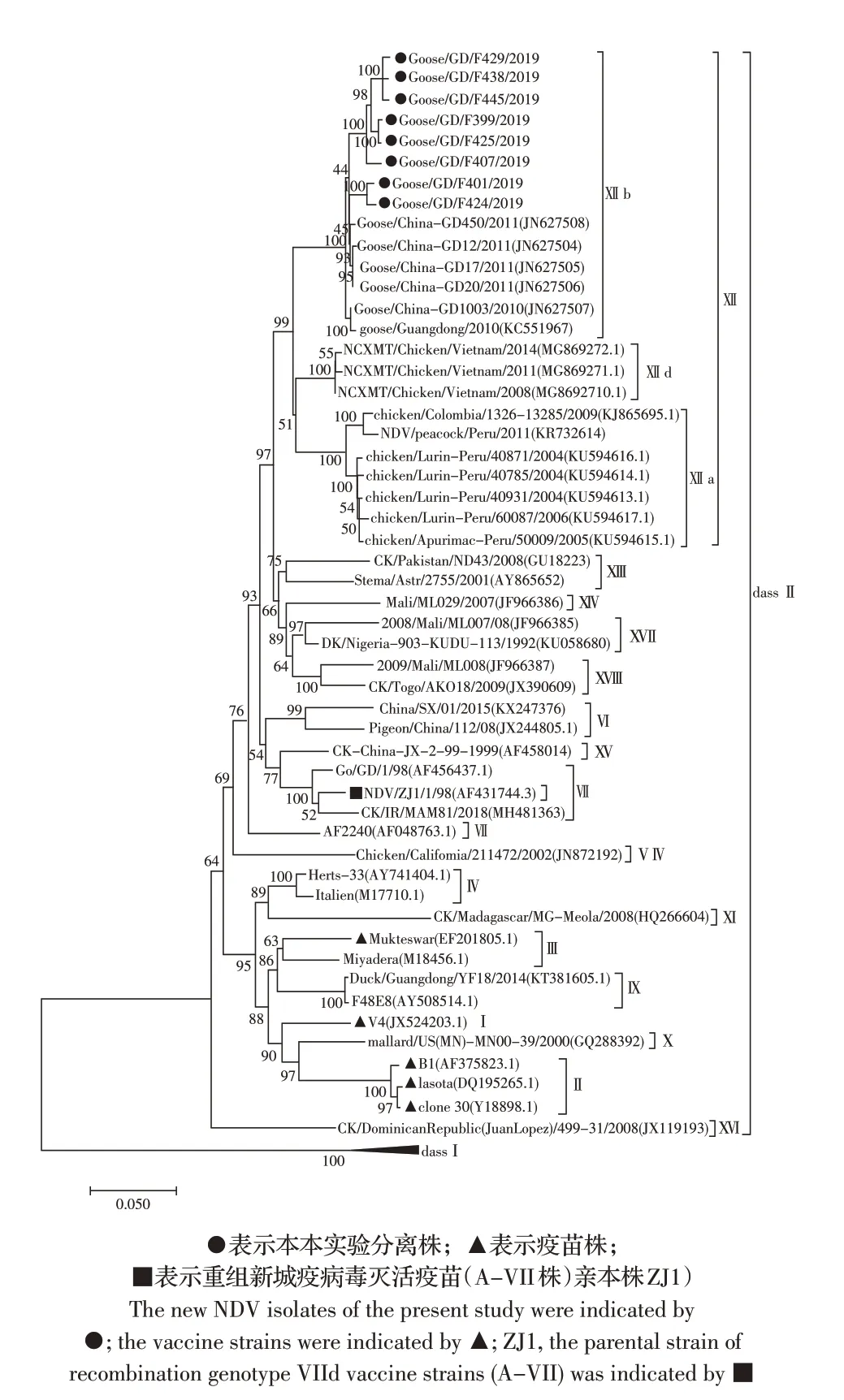
图1 NDV F 基因遗传进化树Fig.1 The phylogenetic tree of NDV F gene

表2 F 蛋白功能位点氨基酸残基变异情况Table 2 Variations of amino acid residues in the functional sites of F protein
