二化螟的多次交配及其对雌蛾产卵量的影响
2020-08-06刘天伟陆明红刘万才郭前爽杜永均
冯 波, 刘天伟, 陆明红, 钟 玲, 郭 荣, 刘万才, 郭前爽, 杜永均,*
(1. 温州医科大学健康与环境生态研究所, 浙江温州 325035; 2. 浙江大学农药与环境毒理研究所, 杭州 310058; 3. 全国农业技术推广服务中心, 北京 100125; 4. 江西省植保植检局, 南昌 330096)
二化螟Chilosuppressalis是我国水稻的重要害虫,目前各地主要依赖化学农药防治。为了解决化学农药防治带来的环境污染和二化螟抗药性快速上升问题,基于性信息素的害虫群集诱杀技术已经在防治二化螟过程中发挥重要作用(左文等, 2008; 杜永均等, 2013)。性诱区的百株卵块数只有对照区的25. 6%~39.8%,卵孵化率下降25.5%,诱蛾量只有对照区的38. 6%,部分地区的诱蛾量甚至下降84.5%,雌雄性比由对照区的1.1上升为诱捕区的3.9,雌虫交配率下降47.7%~54.1%,百株枯心率、枯鞘率和白穗率较对照区分别要低77.9%, 57.0%和44.3%(盛承发等, 2000; 苏建伟等, 2003; Jiaoetal, 2005)。为了更好地理解二化螟的性信息素调控行为,很多学者对二化螟的交配行为进行了研究(焦晓国等, 2006; Quanetal., 2017),分析了日龄(Kanno and Sato, 1978)、温度和湿度(Kanno and Sato, 1979; 余棋等, 2017)、光强度(Kanno, 1981a)、光周期(Kanno, 1981b, 1984)、季节变化(Kanno, 1981c)、寄主差异(Cuong and Cohen, 2003; Ishiguroetal., 2006; Uenoetal., 2006; Huangetal., 2016)等对二化螟交配的影响,为二化螟的群集诱杀和嗅觉适应性提供了大量的理论依据。 王银淑(1989)通过连续3年田间越冬代和连续4代室内饲养二化螟的观察,发现雄蛾多于雌蛾。室内利用稻苗饲养二化螟的雌雄比例甚至达到0.4∶1(雷妍圆等, 2009)。二化螟雄蛾具有多次交配能力,熊兴占等(1984)实验的10头雄蛾全部交配,最多交配6次,平均交配3.8次;焦晓国等(2006)实验的25头雄蛾中有22头交配,最多交配4次,平均交配2.7次。因此很多人认为,性诱剂并不能完全诱杀掉田间的二化螟雄蛾,只能诱杀部分雄蛾,则田间剩余雄蛾可以通过多次交配,与几倍于本身数量的雌蛾交配,完成大量产卵,从而质疑性信息素群集诱杀在二化螟防控中的作用,制约了性信息素群集诱杀技术用于防控具有多次交配习性害虫的大面积推广应用。 为此,本研究调查了二化螟雄蛾的交配次数,比较了不同交配次数雌蛾的繁殖力,为明确性信息素群集诱杀在二化螟防治中的作用奠定了基础。
1 材料与方法
1.1 试虫
2019年3月在浙江省金华市永康市采集含有越冬代二化螟老熟幼虫的稻草并带回浙江大学。实验条件为温度22±2℃,相对湿度70%±5%,光周期13L∶11D。成虫羽化时,每天18∶00时开始,每小时收集一次新羽化的二化螟成虫,直至凌晨1∶00时。根据成虫体型、翅面特征和腹部末端特征鉴别二化螟雌雄。雌雄成虫分开饲养,喂以10%蜂蜜水,每天18∶00时饲喂一次。羽化当天收集的二化螟成虫标记为0日龄,其余日龄以此类推。
1.2 雌雄蛾按1∶1配对时的交配次数
参考焦晓国等(2006)的方法,18∶00时将0日龄未交配雄蛾与1日龄处女雌蛾按1∶1放入350 mL交配杯中,盖上纱布。次日18∶00时解剖与之配对雌蛾的交配囊,检查精包有无,确定雌蛾交配状态。之后放入一头新的1日龄处女雌蛾,持续实验至第6天。根据配对6头雌蛾的交配情况,计算雄蛾的交配次数,同时分析雄蛾的交配动态。雌雄蛾饲养方法同1.1节。本实验共重复5次,每次10组。不同交配次数雄蛾的比例=(不同交配次数雄蛾的数量/10)×100%,某个交配次数雄蛾中进行下一次交配的比例=下一次成功交配的雄蛾数量/当次配对的雄蛾数量×100%。
1.3 雌雄蛾按4∶1配对时的交配次数
考虑到雌雄蛾交配时可能存在选择性,我们设计了如下方法: 18∶00时将0日龄未交配雌雄蛾按4∶1放入350 mL交配杯中,3 d后解剖4头雌蛾交配囊,检查精包有无,确定雌蛾交配状态,并计算雄蛾的交配次数。同时将雄蛾与4头新的0日龄处女雌蛾再次配对,3 d后,解剖4头雌蛾交配囊,检查精包有无,确定雌蛾交配状态。最后根据8头配对雌蛾的交配状况计算雄蛾的交配次数。雌雄蛾饲养方法同1.1节。本实验共重复4次,每次20组。
1.4 雌雄蛾按10∶10配对时的交配次数
18∶00时将1日龄未交配雌雄蛾按10∶10放入交配笼(30 cm×30 cm×30 cm) 中,每日19∶00时开始,利用微弱红光,每隔20 min观察一次雌雄蛾的交配情况。当观察到交配,用50 mL离心管将正在交配的成虫移出,记录交配时间(起始时间、结束时间)。次日18∶00时将交配过1次的存活雄蛾与1日龄处女雌蛾按照10∶10放入交配笼 (30 cm×30 cm×30 cm) 中,从19∶00时开始,按照上述方法观察、移出交配雌雄蛾并记录交配时间。次日18∶00时将交配过2次的存活雄蛾与1日龄处女雌蛾按照10∶10放入交配笼(30 cm×30 cm×30 cm)中,从19∶00时开始,按照上述方法观察、移出交配雌雄蛾并记录交配时间。次日18∶00时将交配过3次的存活雄蛾(不超过10头)与相同数量的1日龄处女雌蛾放入交配笼(30 cm×30 cm×30 cm)中,从19∶00时开始,按照上述方法观察、移出交配雌雄蛾并记录交配时间。雌雄蛾饲养方法同1.1节。1, 2, 3和4次配对分别重复17, 9, 5和3次。
1.5 不同交配次数二化螟雄蛾精巢体积比较
中午13∶00时,解剖1.4节中得到的交配1, 2和3次的5日龄雄蛾精巢,比较不同交配次数雄蛾的精巢体积大小,具体解剖方法如下:将蛾体背面向上,从基部取下腹部,放入75%酒精中,利用镊子从腹基部开始,小心地将腹部各节的背板和腹板去掉,完整地暴露出整个生殖系统,找到精巢,去掉精巢表面的脂肪体,得到完好精巢。摆好精巢姿势,利用Motic显微镜SMZ-168和照相系统Moticam2506(麦克奥迪,厦门)在1×物镜和20×放大倍数条件下拍照。由于二化螟精巢为近圆球体,我们利用软件Motic Plus Images 2.0分别测量了精巢赤道半径a,b(沿着x和y轴)和极半径c的长度,按照椭球体积公式V=4πabc/3计算二化螟精巢体积。各日龄分别解剖雄蛾10头。
1.6 与不同交配次数二化螟雄蛾交配雌蛾的交配囊及精包大小比较
上午9∶00时,解剖1.4节中与不同交配次数二化螟雄蛾进行交配的雌蛾,得到交配囊,在1×物镜和10×放大倍数条件下拍照。之后再解剖精包,同样方法拍照。选择形状为近椭圆体的交配囊和精包,按照1.5节中方法计算交配囊和精包体积。比较与不同交配次数二化螟雄蛾交配雌蛾的交配囊及精包大小。与不同交配次数二化螟雄蛾交配的交配囊和精包数量分别各测量9~14个。
1.7 与不同交配次数二化螟雄蛾交配雌蛾的产卵量比较
将1.4节中与不同交配次数二化螟雄蛾进行交配的雌蛾放入产卵杯中,每日18∶00时检查雌蛾产卵情况,记录产卵量,直至雌蛾死亡,计算雌蛾总产卵量。雌蛾饲养方法同1.1节。与不同交配次数二化螟雄蛾交配的雌蛾各16头。
1.8 数据分析
数据分析采用软件SPSS 17.0。多组平均数间的方差分析采用One-way ANOVA法,显著性分析采用LSD(least significant difference)法,两组平均数间的显著性分析采用Student氏t检验法。利用双变量相关性分析中的Pearson相关系数进行相关性分析,相关性分析的显著性用双尾t检验分析。回归分析采用线性回归法。显著性分析时P>0.05表示差异不显著,P<0.05表示差异显著。
2 结果
2.1 雌雄蛾按1∶1配对时的交配次数
从交配次数分布比例来看,将雌雄蛾按1∶1配对,雄蛾可以交配0~3次(图1: A),其中交配0-3次雄蛾的比例分别为26.0%±2.4%, 38.0%±2.0%, 20.0%±4.5%和16.0%±2.4%(图1: A),雄蛾的交配率为74.0%±2.4%,多次交配雄蛾(交配次数≥2)的比例为36.0%±2.2%。显著性分析表明,交配1次的雄蛾比例显著高于交配0, 2和3次的(P≤0.005),交配0次的雄蛾比例显著高于交配3次的(P=0.02)。 交配雄蛾平均交配1.7±0.1次。
从某一交配频次雄蛾进行下一次交配的比例统计来看,1∶1配对雄蛾中有74.0%±2.4%会进行交配(图1: B);在交配1次的雄蛾中,有48.6%±2.7%会进行第2次交配;在交配2次的雄蛾中,有46.7%±9.4%会进行第3次交配。雄蛾进行第1次交配的比例显著高于交配第2和3次的(P≤0.041)(图1: B)。
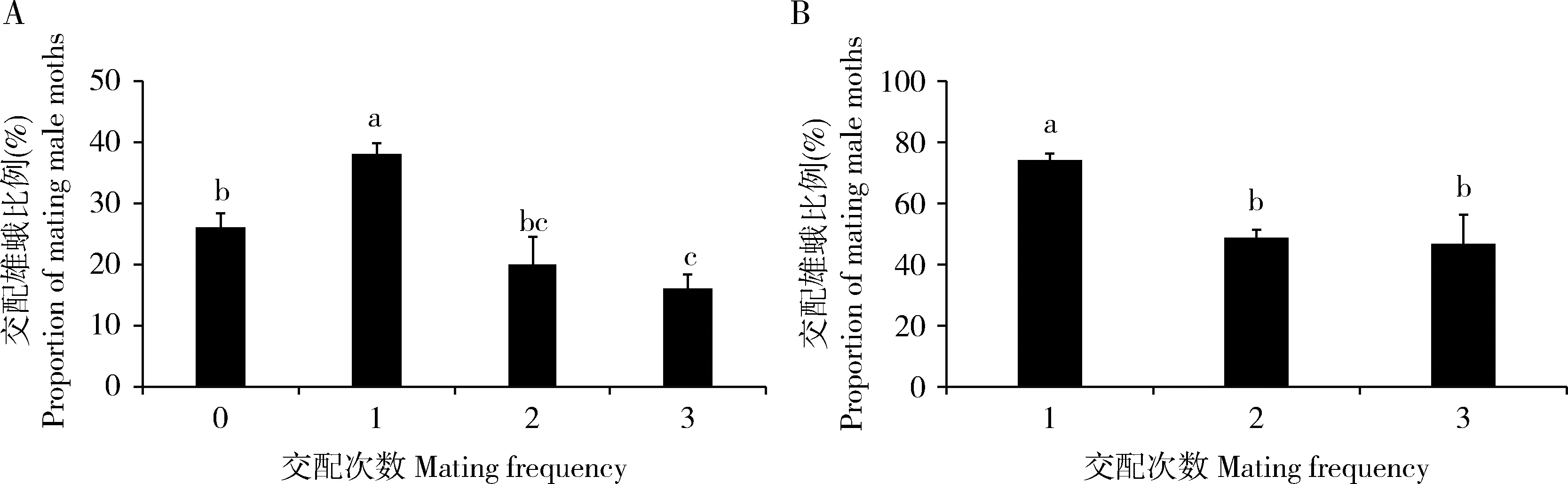
图1 雌雄按1∶1配对时二化螟雄蛾的交配次数Fig. 1 Mating frequency of male moths of Chilo suppressalis when paired in the ratio of 1 female to 1 maleA: 不同交配次数雄蛾的比例Proportion of male moths with different mating frequencies; B: 某个交配次数雄蛾中进行下一次交配的比例Proportion of male moths taking the next mating at a certain mating frequency. 柱上不同字母表示不同交配次数雄蛾的交配比例差异显著(LSD法, P<0.05)。Different letters above bars indicate significant differences in the proportion of mating male moths among different mating frequencies by LSD method (P<0.05). 图2和3同The same for Figs. 2 and 3.
从交配雄蛾的日龄分布来看,0-5日龄雄蛾分别交配了14, 21, 13, 8, 5和2头,其中1日龄和2日龄交配雄蛾中分别有16和7头雄蛾为首次交配,从3日龄雄蛾开始,新交配数量极少;有5, 4和1头雄蛾分别在0-1日龄、1-2日龄和4-5日龄连续交配,有1头雄蛾在0-2日龄连续交配;在进行多次交配的18头雄蛾中,分别有8, 8和2头雄蛾分别在0-2日龄首次交配,表明多次交配雄蛾绝大部分来自于0-1日龄交配雄蛾(表1)。

表1 二化螟雄蛾的交配动态Table 1 Mating dynamics of male moths of Chilo suppressalis
2.2 二化螟雌雄蛾按4∶1配对时的交配次数
从交配次数分布比例来看,雌雄蛾按4∶1配对,雄蛾可以交配0~4次(n=80),其中交配0-4次的雄蛾比例分别为30.6%±2.6%, 19.5%±2.1%, 25.6%±2.1%, 19.4%±1.6%和4.9%±2%(图2: A),雄蛾的交配率为69.4%±2.6%,多次交配雄蛾的比例为51.3%±2.3%。雌雄蛾按4∶1配对时,交配雄蛾的平均交配次数为2.1±0.1次,显著高于雌雄蛾按1∶1配对时雄蛾的平均交配次数(P=0.02)。显著性分析表明,交配0次的雄蛾比例显著高于交配1次和3次的(P≤0.016),再显著高于交配4次的(P≤0.002)(图2: A)。
从某一交配频次雄蛾进行下一次交配的比例统计来看,雌雄蛾按4∶1配对,雄蛾中有69.4%±2.6%会进行交配(图2: B);在交配1次的雄蛾中,有72.0%±2.4%会进行第2次交配;在交配2次的雄蛾中,有48.9%±3.2%会进行第3次交配;在交配3次的雄蛾中,有19.2%±8.2%会进行第4次交配。雄蛾进行第3次交配的比例显著低于交配第1和2次的(P≤0.003),进行第4次交配的雄蛾比例显著低于交配第3次的(P=0.015)(图2: B)。在交配1次的雄蛾中,雌雄蛾按4∶1配对时,进行第2次交配的雄蛾比例显著高于雌雄蛾按1∶1配对时的雄蛾比例(P<0.001)。

图2 雌雄按4∶1配对时二化螟雄蛾的交配次数Fig. 2 Mating frequency of male moths of Chilo suppressalis when paired in the ratio of 4 females to 1 maleA: 不同交配次数雄蛾的比例Proportion of male moths with different mating frequencies; B: 某个交配次数雄蛾中进行下一次交配的比例Proportion of male moths taking the next mating at a certain mating frequency.
2.3 二化螟雌雄蛾按10∶10配对时的交配次数
从交配次数分布比例来看,雌雄蛾按照10∶10配对时,雄蛾可以交配0~4次,其中交配0-4次的雄蛾比例分别为34.5%±2.4%, 27.8%±2.7%, 20.7%±2.1%, 10.4%±1%和6.7%±1.6%(图3: A)。交配雄蛾平均交配1.9±0.1次。显著性分析表明,交配0次的雄蛾比例显著高于交配2次的(P=0.001),再显著高于交配3和4次的(P≤0.002)(图3: A)。
从某一交配频次雄蛾进行下一次交配的比例统计来看,雌雄蛾按10∶10配对,雄蛾中有65.5%±2.4%会进行交配;在交配1次的雄蛾中,有57.3%±4.3%会进行第2次交配;在交配2次的雄蛾中,有44.8%±3.3%会进行第3次交配;在交配3次雄蛾中,有36.9%±4.2%会进行第4次交配。雄蛾进行第1和2次交配的比例显著高于交配第3和4次的(P≤0.040)。多次交配雄蛾的比例为37.8%±2.5%(图3: B)。

图3 雌雄按10∶10配对时二化螟雄蛾的交配次数Fig. 3 Mating frequency of male moths of Chilo suppressalis when paired in the ratio of 10 females to 10 malesA: 不同交配次数雄蛾的比例Proportion of male moths with different mating frequencies; B: 某个交配次数雄蛾中进行下一次交配的比例Proportion of male moths taking the next mating at a certain mating frequency.
2.4 不同交配次数雄蛾的交配持续时间比较
从交配持续时间来看,第1, 2和3次交配的持续时间分别为106.1±3.9, 98.8±3.6和145.4±9.3 min,其中第3次交配的持续时间显著长于第1和2次交配(P<0.001),第1和2次交配的持续时间无显著差异(P=0.173)(图4)。

图4 不同交配次数二化螟雄蛾的交配持续时间Fig. 4 Mating duration of male moths of Chilo suppressalis with different mating frequencies 柱上不同字母表示差异显著(LSD法, P<0.05)。Different letters above bars indicate significant differences by LSD method (P<0.05). 图5和7同The same forFigs. 5 and 7.
2.5 不同交配次数雄蛾精巢大小比较
从雄蛾精巢大小来看,交配1, 2和3次雄蛾的精巢体积分别为0.187±0.022, 0.193±0.021和0.187±0.026 mm3(图5),显著性分析表明交配1-3次雄蛾的精巢体积之间差异不显著(P≥0.857)。

图5 不同交配次数二化螟雄蛾的精巢体积Fig. 5 The testicular volume of male moths of Chilo suppressalis with different mating frequencies
2.6 与不同交配次数雄蛾交配雌蛾的交配囊和精包比较
从雌蛾交配囊形状来看,新交配二化螟雌蛾的交配囊表现出不同形状,包括肾形、椭圆形和倒锥形(图6: A, B)。二化螟的交配囊为膜质,撕开交配囊后,发现其内含有的精包形状和交配囊形状相似(图6: D, E),表明交配后雌蛾交配囊的形状主要由其内部的精包形状决定。从雌蛾交配囊体积来看,与第1和2次交配雄蛾进行交配雌蛾的交配囊体积分别为16.6±1.0和20.3±2.5 mm3(图6: C),其内含的精包体积分别为12.8±2.0和13.6±1.7 mm3(图6: F)。与第1和2次交配雄蛾进行交配雌蛾的交配囊和精包体积均差异不显著(P≥0.198)。

图6 刚完成交配二化螟雌蛾的交配囊(A-C)和精包(D-F)Fig. 6 The bursa copulatrix (A-C) and spermatophore (D-F) of the newly mated female moths of Chilo suppressalis A, B: 刚完成交配雌蛾中不同形状的交配囊Bursa copulatrix of the newly mated female; C: 与不同交配次数雄蛾完成交配后雌蛾的交配囊体积Volume of bursa copulatrix of the female moths mated with male moths at different mating frequencies; D-E: 刚完成交配雌蛾交配囊中不同形状的精包Spermatophore in the bursa copulatrix of the newly mated female; F: 与不同交配次数雄蛾完成交配后雌蛾交配囊中的精包体积Volume of spermatophore in the bursa copulatrix of the female moths mated with male moths at different mating frequencies. ns: 无显著差异(Student氏t检验, P>0.05) No significant difference (Student’s t-test, P>0.05).
2.7 与不同交配次数雄蛾交配雌蛾的产卵量比较
从雌蛾产卵量来看,与第1, 2和3次交配雄蛾进行交配的雌蛾的产卵量分别为77.9±16.5, 79.7±18.2和77.5±18.7枚,各产卵量之间几乎无差异(P≥0.936)(图7)。

图7 二化螟雄蛾交配次数对雌蛾产卵量的影响Fig. 7 Effect of mating frequency of male moths of Chilo suppressalis on the number of eggs laid by female
3 讨论
3.1 二化螟雌雄蛾交配的选择性导致部分雄蛾在交配前死亡
在3种雌雄配对方式中,分别有26.0%, 30.6%和34.5%的雄蛾在交配之前即死亡。雄蛾在交配前死亡的情况在焦晓国等(2006)的研究中也有发现。表明并不是所有雄蛾均能够进行交配。事实上,二化螟雌蛾的交配率也不会达到100%。王银淑(1989)从室内大量二化螟交配雌蛾中随机选择50头进行解剖,连续解剖3代,发现交配0次的比例为36.5%。焦晓国等(2006)用25头雌蛾进行交配实验时,有2头在交配前死亡。我们还发现,雌雄蛾按4∶1配对时的雄蛾交配次数显著高于按1∶1配对时(P=0.02),这是由于在交配1次雄蛾中,按4∶1配对时雄蛾进行第2次交配的比例显著高于按1∶1配对时(P<0.001)。上述结果均表明二化螟雌雄蛾在交配时会对交配对象进行选择。同一日龄雄蛾中,已交配雄蛾的精巢体积显著大于未交配雄蛾(冯波等, 2019),说明二化螟雌蛾会选择精巢体积更大的雄蛾进行交配,交配是雌雄双方选择的结果,交配选择可能是为了使种群具有更强的竞争能力。但是雌雄蛾以10∶10配对时雄蛾的平均交配次数与雌雄蛾以1∶1和4∶1配对时雄蛾的平均交配次数均无显著差异(P≥0.152)(图1~3)。同时通过调整光周期使茭白二化螟和水稻二化螟的交配时间重叠,水稻二化螟雌(雄)蛾并不会明显选择水稻二化螟雄(雌)蛾进行交配(Quanetal., 2017)。
3.2 具有多次交配能力的0-1日龄强壮雄蛾在首次交配前被合成性信息素诱捕
合成性信息素引诱的田间二化螟雄蛾精巢小于0-1日龄未交配雄蛾,大于2-6日龄未交配雄蛾(冯波等, 2019),同时性诱雄蛾附腺端部内含物等级显著高于2-6日龄未交配雄蛾,第3和4段非角质化区内含物等级仅显著低于刚羽化(0日龄)雄蛾(郭前爽等, 2019)。考虑到性信息素引诱雄蛾的解剖时间延后了12 h,推测田间性信息素引诱雄蛾中大部分是0-1日龄。室内试验还证实,被性信息素诱捕雄蛾的精巢体积显著大于未被性信息素诱捕雄蛾(冯波等, 2019)。本研究发现二化螟雄蛾的首次交配主要发生在0-1日龄,从3日龄开始,雄蛾进行首次交配的比例极低,同时能够连续2晚进行交配的雄蛾比例较低,0-1日龄交配雄蛾中的绝大部分为首次交配(表1)。这和雄蛾生殖系统的解剖结果相似,交配后24 h,已交配雄蛾生殖器内含物等级虽然和未交配雄蛾无显著差异,但是直到交配后36 h,二化螟雄蛾生殖器内含物等级才恢复到未交配状态(郭前爽等, 2019)。因此田间性诱剂引诱的绝大部分雄蛾应该为未交配的0-1日龄蛾。本研究还发现,在进行多次交配雄蛾中,0和1日龄首次交配雄蛾的比例分别为44.4%,表明被性信息素引诱的田间二化螟雄蛾中很多个体都具有多次交配能力,未被性信息素引诱雄蛾中很少个体能够进行多次交配。这主要是由于雄蛾精巢体积随日龄的增加而变小,已交配雄蛾的精巢体积显著大于未交配雄蛾,交配并不影响雄蛾精巢的发育(冯波等, 2019),因此0和1日龄首次交配雄蛾的精巢很大,大于其余日龄首次交配雄蛾,从而保证了这些雄蛾进行多次交配的能力。精巢体积大小和交配次数的正相关性在其他多种昆虫中得到证实,例如精巢大的雄蝶可能具有更多的交配次数(Gage, 1994),交配1次的四纹豆象Callosobruchusmaculatus种群比多次交配种群的精巢体积更小(Gayetal., 2009)。我们利用3种配对方式(雌雄蛾按1∶1, 4∶1和10∶10配对)对二化螟雄蛾的交配次数进行了详细研究,发现二化螟雄蛾确实可以进行多次交配,但是能够进行多次交配的雄蛾比例并不太高,分别为36.0%, 51.3%和37.8%(图1-3)。
3.3 性信息素群集诱杀防控二化螟的有效性
二化螟雄蛾先于雌蛾羽化,因为雄蛾的发育历期为38.8 d,雌蛾的发育历期为40.6 d(肖丹凤和胡阳, 2010)。雄蛾羽化后即可交配,羽化当晚的交配率为27.9%±2.7%,1日龄雄蛾的交配率达66.7%±5.3%(冯波等, 2019),可以在交配窗口期利用性诱剂将先羽化的雄蛾诱杀。性信息素能够诱捕大量具有多次交配能力的0-1日龄强壮雄蛾,并且这种诱捕是在其与雌蛾进行首次交配前完成。将这部分雄蛾诱杀后,虽然田间还有一定数量的雄蛾,但是这些剩余雄蛾中有很多不会进行交配;即使进行交配,它们也仅能进行1次交配,很难进行多次交配,因此性信息素群集诱杀能够急剧降低田间雌蛾的交配率。虽然大部分二化螟雌蛾一生只交配1次,但是仍然有部分雌蛾可以交配2次或更多次(熊兴占等, 1984; 孙丽娟等, 2002),这也会在一定程度上降低了田间交配雌蛾的数量。由于性信息素将精巢体积更大的雄蛾诱杀,田间剩余雄蛾的精巢较小,产生的精子数量较少,交配成功后雌蛾产下的受精卵的数量也较少。因此正确使用性诱剂可以大量诱杀交配能力强的二化螟雄性个体,降低田间雄蛾数量,极大地降低田间雄蛾的交配能力和雌蛾的交配率和产卵量,有效压低田间二化螟种群数量,取得很好的性诱防控效果。但是二化螟群集诱杀的防控效果极大地取决于性诱剂的引诱效果。如果性诱剂的引诱力不强,不能与田间雌蛾形成有效竞争,例如早期的橡皮头诱芯;性诱捕器的诱杀效率不高,例如水盆诱捕器,则性诱剂诱杀的雄蛾数量少,很多可能是已经交配过的雄蛾(Huetal., 2018),性诱防效可能就较差甚至没有。
3.4 二化螟雄蛾的交配次数对雌蛾产卵量无显著影响
虽然雄蛾进行第3次交配的持续时间显著长于第1和2次交配,但是交配1-3次雄蛾的精巢体积之间差异不显著;与第1和2交配雄蛾进行交配后,雌蛾交配囊和内含的精包体积均差异不显著;和第1, 2和3次交配雄蛾进行交配后,雌蛾的产卵量之间几乎无差异(图4~7),这些结果表明雄蛾的交配次数对与之交配雌蛾的繁殖力没有显著影响。熊兴占等(1984)也发现与第1-4次交配雄蛾交配雌蛾的产卵量和卵孵化率之间无明显差异。焦晓国等(2006)认为雄蛾不同交配史对雌蛾产卵期、产卵量、卵孵化率和寿命等生物学指标没有显著不良影响。另外,焦晓国等(2006)还认为雄蛾交配日龄对雌蛾生殖力、寿命、产卵量、产卵期等都没有显著影响。
本研究中未交配雄蛾的比例高于熊兴占等(1984)和焦晓国等(2006),而平均交配次数低于熊兴占等(1984)和焦晓国等(2006)。造成这种差异的原因可能包含如下两方面:一方面是本研究使用的雄蛾数量较大,约为300头,而熊兴占等(1984)和焦晓国等(2006)等使用的雄蛾数量较小,仅10~25头;另一方面是实验方法存在差异。实验方法差异具体分为以下几点:(1)本研究虫源来自浙江金华,熊兴占等(1984)的虫源来自重庆,焦晓国等(2006)的虫源未知。(2)熊兴占等(1984)在实验中放入了秧苗,本研究为了便于观察和抓捕交配中的二化螟,在实验过程中并未放入秧苗。有研究证实寄主植物可以促进二化螟成虫的交配 (Samudraetal., 2002)。(3)温度、湿度、光周期等实验条件存在差异。焦晓国等(2006)的实验条件为温度25℃和光周期16L∶8D,熊兴占等(1984)在温度25℃和90%相对湿度条件下实验,本研究则为温度22±2℃,相对湿度70%±5%,光周期13L∶11D。研究表明温度、湿度、光周期等均会影响二化螟的交配行为(Kanno and Sato, 1979; Kanno, 1981b, 1981c, 1984)。事实上,研究结果出现差异在二化螟中是一个普遍现象,例如Konno和Tanaka (1996)认为水稻二化螟的交配行为发生在黑暗第2-5小时,但是Samudra 等(2002)却认为水稻二化螟雌蛾几乎在整个黑暗期(黑暗第2-9小时)都有求偶行为,这其中一个原因可能与季节有关。同一研究者在不同时间发表结果也差异很大,例如Konno和Tanaka (1996)认为水稻二化螟和茭白二化螟1日龄蛾的交配率分别为56.7%和65.2%,但是Kanno (1998) 后来认为水稻二化螟和茭白二化螟1日龄蛾的交配率分别为80%和20%。因此我们并不能将不同的研究结果进行简单比较。
