茶足柄瘤蚜茧蜂滞育和非滞育蛹中与能量代谢相关的差异表达蛋白
2020-08-06韩海斌刘爱萍高书晶徐林波岳方正黄海广
刘 敏, 韩海斌, 刘爱萍,*, 高书晶, 徐林波, 岳方正, 黄海广
(1. 中国农业科学院草原研究所, 呼和浩特 010000; 2. 国家林业和草原局森林和草原病虫害防治总站, 沈阳 110034; 3. 内蒙古自治区林业科学研究院, 呼和浩特 010000)
茶足柄瘤蚜茧蜂Lysiphlebustestaceipes是一种寄生性天敌,寄主蚜虫种类广泛,包括危害经济作物的麦二叉蚜Schizaphisgraminum、棉蚜Aphisgossypii、玉米蚜Rhopalosiphummaidis和大豆蚜Aphisgylcnies以及豆科牧草和沙生植物的苜蓿蚜Aphiscraccivora等(Rodrigues and Bueno, 2001; Silvaetal., 2008; 刘兴龙等, 2009)。滞育是指昆虫在不利环境条件下受到某些信号的刺激,通过体内一系列生理生化变化,使其生长、发育和繁殖暂时停止的生命现象,是对不利环境的遗传性适应(Tauberetal., 1986; Saunders, 2002),滞育一旦开始,不会因为环境条件的转变而立即解除滞育,通常会持续一段时间(刘流等, 2010)。滞育不仅可以帮助昆虫度过不良环境,维持个体的生存;而且可以使昆虫种群发育整齐,提高雌雄个体间的交配机率,保证种群的繁衍(王满囷和李周直, 2004)。利用茶足柄瘤蚜茧蜂滞育特性来延长其产品的货架期,可以更好地防治害虫。目前国内外对寄生蜂滞育的研究主要包括诱导滞育的条件、滞育特征、滞育的亲代效应、滞育后发育、滞育的分子机制等(张洪志等, 2018)。随着基因组学、转录组学、蛋白质组学、代谢组学等生物技术近年来的快速发展与广泛应用,昆虫滞育的机理正逐步被揭示。滞育期间,昆虫的血淋巴或脂肪体中存在某些蛋白质,且浓度较高,随着滞育的终止,这些蛋白质逐渐消失,而在非滞育昆虫中无此类蛋白质(或仅有极微量痕迹),它们被认为与昆虫滞育的发生有关,称为滞育关联蛋白(diapause-associated protein, DAP)。昆虫在滞育过程中,代谢速率缓慢,形态上无变化,也无组织分化和器官发育,但生理过程仍继续进行,如神经分泌、脂类代谢及糖类代谢等(Duboisetal., 1956; Haykawa and Chino, 1981; Puirouxetal., 1989)。昆虫能够在极端环境条件下成功存活正是由于这种特殊的生理代谢机制(Mansingh, 1971; Storey and Storey, 1988; Denlinger, 1991)。
能量代谢是有机体在物质代谢过程中所伴随能量释放、转移、贮存与利用的过程,昆虫利用这些能量来维持生命活动。为探究昆虫滞育过程中能量代谢分子机制,本研究根据实验室前期研究基础,对茶足柄瘤蚜茧蜂进行滞育诱导,采用iTRAQ技术比较茶足柄瘤蚜茧蜂滞育组与非滞育组的蛋白含量,并结合生物信息学方法揭示了茶足柄瘤蚜茧蜂滞育期间蛋白的变化规律,筛选出与能量代谢相关的滞育关联蛋白并分析功能,为深入分析茶足柄瘤蚜茧蜂滞育机理提供参考。 这一工作对研究昆虫滞育分子机理与应用天敌防治害虫具有重要意义,同时也可为滞育后害虫发生量与发生期预测预报提供一种依据。
1 材料与方法
1.1 试虫来源和样品收集
1.1.1试虫来源:载体植物蚕豆Viciafaba苗为室内培育,寄主苜蓿蚜采自中国农业科学院草原研究所沙尔沁基地的羊柴Hedysarummongolicum植株上,将苜蓿蚜转接在温室内的水培蚕豆苗上繁殖,确保接在蚕豆苗上的苜蓿蚜未被天敌寄生,接虫后对蚕豆苗进行笼罩(100目防虫网),在温室内饲养5代以上作为供试寄主虫源。从中国农业科学院草原研究所沙尔沁基地的羊柴植株上采集被寄生的苜蓿蚜僵蚜,从中挑取茶足柄瘤蚜茧蜂未羽化破壳的僵蚜置于人工气候箱温度(25±1)℃,相对湿度(70±1)%,光周期14L∶10D条件下培养,挑选羽化后茶足柄瘤蚜茧蜂转移至试管(10 cm×3 cm)内,用20%的蜂蜜水作为补充营养,接入上述实验室条件下培养的苜蓿蚜2-3龄若蚜上,在室温下养虫笼中按照足柄瘤蚜茧蜂∶苜蓿蚜=1∶100释放成对刚羽化茶足柄瘤蚜茧蜂和苜蓿蚜,建立茶足柄瘤蚜茧蜂种群作为供试虫源。 茶足柄瘤蚜茧蜂在室温下用苜蓿蚜有效扩繁10代以上,取羽化24 h内的成蜂待用。
1.1.2样品收集:根据实验室前期研究基础可知,苜蓿蚜若蚜被茶足柄瘤蚜茧蜂寄生后,寄生蜂卵继续发育120 h,此时僵蚜体内寄生蜂处于高龄幼虫(3-4龄)阶段,高龄幼虫为茶足柄瘤蚜茧蜂感受滞育信号的敏感虫态,将此时的僵蚜放入人工气候箱中进行滞育诱导。高龄幼虫处于滞育环境条件时,并不会立刻停止发育,而是继续发育一段时间,经实验验证,当发育至蛹时,便不再继续发育(孙程鹏, 2018)。本研究中,基于1.1.1节中供试虫源,诱导茶足柄瘤蚜茧蜂滞育的温光组合为温度8℃和光周期8L∶16D,诱导时长为30 d。选取经过30 d滞育诱导的僵蚜300头,解剖出茶足柄瘤蚜茧蜂活蛹, 50头/管放入液氮中速冻暂时保存,然后作为滞育组样品放入-80℃冰箱中保存;苜蓿蚜若蚜被茶足柄瘤蚜茧蜂寄生后,放置在(25±0.5)℃、RH (70±5)%、光周期14L∶10D、光照强度8 800 lx(人工气候箱,上海一恒公司MGC-HP系列)条件下,寄生蜂卵继续发育168 h(此时蚜茧蜂处于蛹态),对300头僵蚜进行解剖,挑选饱满、有活力的茶足柄瘤蚜茧蜂蛹, 50头/管放入液氮中速冻暂时保存,然后作为非滞育组样品放入-80℃冰箱中保存。
1.2 滞育组与非滞育组茶足柄瘤蚜茧蜂蛋白提取
蛋白质提取采用裂解液法(徐珊珊等, 2008)。采用考马斯亮蓝法(Bradford 法)(胡文霞,2011)测定蛋白浓度。取100 μg蛋白样品,加入适量蛋白溶解液(8 mol/L尿素, 100 mmol/L TEAB, pH 8.5)补足体积至100 μL,胰蛋白酶(1 μg/μL) 2 μL和TEAB缓冲液(100 mmol/L) 500 μL,混匀后于37℃酶切过夜,取上清通过C18除盐柱进行脱盐处理,然后加入足量iTRAQ标记试剂(溶于异丙醇),室温下颠倒混匀反应1 h,取等体积标记后的蛋白样品混合,除盐后冻干。配制流动相A液(2%乙腈和98%水,氨水调至pH 10)和B液(98%乙腈和2%水,氨水调至pH 10)。使用1 mL A液溶解标记后的混合样品粉末,离心后取1 mL上清进样。经过脱盐处理, 真空干燥,每分钟收集1管,冻干后各加入0.1%甲酸溶解,经高效液相色谱 (HPLC) 预分离。配制流动相A液(100%水和0.1%甲酸)和B液(80%乙腈和0.1%甲酸)。对收得馏分上清各取2 μg样品进样,液质检测。使用Q ExactiveTMHF-X质谱仪,进行二级质谱检测。
1.3 数据分析
使用软件Discoverer2.2对生成质谱检测的原始数据(.raw)进行查库鉴定及定量分析。Discoverer2.2软件对检索结果做了进一步过滤以提高分析结果质量,降低假阳性率,可信度在95%以上的肽段匹配谱图(peptide spectrum matches, PSMs)为可信PSMs,至少包含一个特有肽段的蛋白为可信蛋白,只保留可信的PSMs和蛋白,经错误发现率(false discovery rate, FDR)验证去除FDR>5%的肽段和蛋白。将每个蛋白在滞育组和非滞育组中的所有生物学重复定量值均值的比值作为差异倍数(fold change, FC)。为了判断差异的显著性,将每个蛋白在滞育组和非滞育组样品中的相对定量值进行了T检验,并计算相应的P值,以此作为显著性指标。当FC≥2.0,同时P≤0.05时,蛋白表现为表达量上调;当FC≤0.50,同时P≤0.05时,蛋白表现为表达量下调。
利用GO功能显著性富集分析将滞育组和非滞育组样品差异表达蛋白(differentially expressed proteins, DEPs)向Gene Ontology数据库(http:∥www.geneontology.org/)的各个term映射,计算每个term的蛋白质数目,然后应用超几何检验找出与所有蛋白质背景相比,在DEPs中显著富集的GO条目(term)。其计算公式:
其中N为所有蛋白中具有GO注释信息的蛋白数目,n为N中差异蛋白的数目,M为所有蛋白中注释到某个GO条目的蛋白数目,X为注释到某个GO条目的差异蛋白数目。计算得到P值,以P≤0.05为阈值,满足此条件的GO term定义为在差异蛋白质中显著富集的GO term。通过GO显著性分析能确定DEPs行使的主要生物学功能。
利用KEGG pathway显著性富集分析方法对滞育组和非滞育组样品DEPs进行功能富集分析,确定DEPs参与的最主要生化代谢途径和信号转导途径。
2 结果
2.1 茶足柄瘤蚜茧蜂滞育与非滞育蛹中差异表达蛋白
本研究中定量蛋白总数为7 251个,根据1.3节中筛选差异蛋白的条件,对定量到的蛋白进行筛选,滞育组与非滞育组中DEPs有135个,滞育组上调表达蛋白有38个(FC≥2.0,P≤0.05),上调表达量最高的蛋白为茶足柄瘤蚜茧蜂knottin样蛋白(序列ID为Cluster-35028.0,FC=4.616);下调表达蛋白有97个(FC≤0.50,P≤0.05),下调表达量最高的蛋白为大蜜蜂Apisdorsata不均一核糖核蛋白(heterogeneous nuclear ribonucleoprotein) M-like (序列ID为Cluster-50256.0,FC=0.145)。火山图(图1)可以非常直观地展现出滞育组与非滞育组样本间的差异蛋白,及其统计学显著性(P值)和表达水平差异程度(FC)。

图1 茶足柄瘤蚜茧蜂滞育与非滞育蛹差异表达蛋白火山图Fig. 1 Volcano plot of differentially expressed proteins (DEPs) in diapause and non-diapause pupae of Lysiphlebus testaceipes None: 表达差异不显著的蛋白Proteins without significant expression difference; Up: 上调DEPs (Up-regulated DEPs); Dwon: 下调DEPs (Down-regulated DEPs). 图2同The same forFig.2. 横坐标表示滞育蛹(D)与非滞育蛹(ND)间差异蛋白的差异倍数(log2值),纵轴表示P值(-log10值)。Transverse coordinate indicates the fold change (log2P-value) of DEPs in diapause pupae (D) and non-diapause pupae (ND), and longitudinal axis represents the P-value (-log10 value).
2.2 茶足柄瘤蚜茧蜂滞育与非滞育蛹中差异表达蛋白的GO富集分析
图2中,GO注释到的差异蛋白数目为90,分为生物学过程、细胞成分和分子功能3类,富集到154条条目,共有44个GO条目显著富集;在生物学过程中参与有机物代谢过程的蛋白数最多,高分子代谢过程和蛋白质代谢过程次之;在细胞成分中与胞内细胞器和细胞质功能相关的蛋白数较多;在分子功能部分中参与结构分子活性和核糖体结构组成的蛋白质数量较多。与天冬氨酸转运(aspartate transport)、L-谷氨酸转运(L-glutamate transport)、胆碱脱氢酶活性(choline dehydrogenase activity)、胆碱生物合成甘氨酸甜菜碱(glycine betaine biosynthetic process from choline)等条目相关的蛋白质在滞育阶段显著上调表达。

图2 茶足柄瘤蚜茧蜂滞育与非滞育蛹间差异表达蛋白的GO功能富集Fig. 2 GO function enrichment of differentially expressed proteins in diapause and non-diapause pupae of Lysiphlebus testaceipes
2.3 茶足柄瘤蚜茧蜂滞育与非滞育蛹中差异表达蛋白的KEGG富集分析
KEGG注释到64个差异蛋白,共富集到97条KEGG pathway,对富集通路进行显著性分析发现,除与人类疾病相关的通路外,有3条途径显著富集到KEGG pathway上,分别是核糖体(ribosome)、氧化磷酸化(oxidative phosphorylation)和逆行内源性大麻素信号(retrograde endocannabinoid signaling)。而富集到这些条目及通路中的蛋白质与能量代谢及抗逆性有密切关系。根据富集结果,绘制富集到的KEGG通路的气泡图(图3, 只展示top20的结果)。
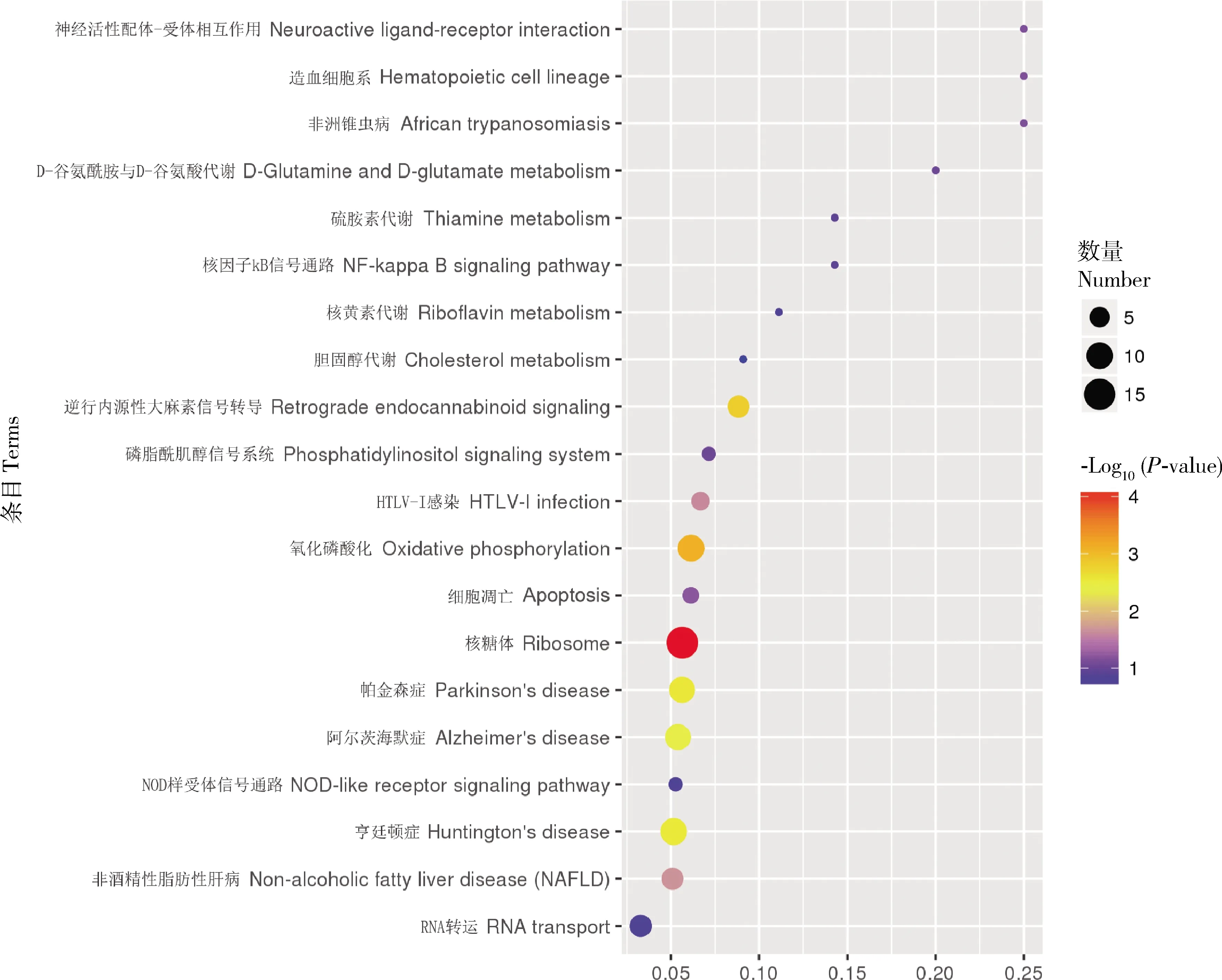
图3 茶足柄瘤蚜茧蜂滞育蛹与非滞育蛹差异表达蛋白KEGG富集气泡图Fig. 3 Scatter plot of KEGG enrichment of differentially expressed proteins (DEPs) in diapause and non-diapause pupae of Lysiphlebus testaceipes 图中横坐标轴为相应通路中DEPs的数目与鉴定出的总蛋白数目的比值,值越大,说明在该通路中DEPs富集程度越高;圆点的颜色代表超几何检验的P值,值越小,说明检验的可靠性越大、越具统计学意义。点的大小代表相应通路中DEPs的数目,值越大,该通路内DEPs越多。The abscissa in theFigure is the ratio of the number of DEPs in the corresponding pathway to the number of total proteins identified. The larger the value is, the higher the concentration of DEPs in this pathway is. The color of dots represents the P-value of hypergeometric test. The smaller the value is, the more reliable and statistically significant the test is. The size of the dots represents the number of DEPs in the corresponding pathway. The larger the dot, the more DEPs in the pathway.
3 讨论
近年来,发现了大量滞育关联蛋白,并鉴定分析了一些蛋白。例如,Hao等(2012)对菜蛾盘绒茧蜂Cotesiavestalis进行了滞育研究,主要研究了过氧化物相关酶在其亲代效应中的作用,结果显示,亲代的滞育蛹过氧化物酶活性升高,过氧化氢酶活性降低,但是在滞育子代的卵中过氧化物酶和过氧化氢酶的活性显著提高,并以此推测在代际间过氧化氢具有进行信号传导的作用;Colinet等(2012)利用2D-DIGE技术对滞育与非滞育翼蚜外茧蜂Praonvolucre的蛋白进行检测,确定了221个差异表达显著的蛋白,对这些蛋白利用质谱技术鉴定,最终鉴定出细胞骨架蛋白、ATP 结合蛋白、角质层类蛋白、应激蛋白、糖酵解、脂代谢、蛋白质代谢等重要代谢过程中的酶等;黄凤霞等(2015)利用iTRAQ技术,对滞育与非滞育烟蚜茧蜂Aphidiusgifuensis进行蛋白检测,在滞育阶段发现278个蛋白上调表达。本研究中下调表达量最高的蛋白为大蜜蜂不均一核糖核蛋白 M-like(序列ID为Cluster-50256.0,FC=0.145),该蛋白被用作转录调节剂,促进转录抑制(黄嘉等, 2004)。这些差异表达蛋白主要与糖代谢、脂代谢、蛋白质代谢等代谢过程及氨基酸转运、能量产生与转化,各种代谢酶等有关。能量代谢是滞育昆虫成活的关键,滞育期间的营养储备水平直接影响昆虫的存活情况以及滞育后的发育和生殖(张倩等, 2019)。KEGG通路分析显示,茶足柄瘤蚜茧蜂蛹滞育相关蛋白在氧化磷酸化通路显著上调。在有氧条件下,氧化磷酸化作用是需氧细胞生物生命活动的主要能量来源,在细胞内的有机分子经氧化分解形成CO2和H2O,并释放出能量使ADP和Pi合成ATP(王镜岩等, 2008)。其中有10个与能量产生及转化有关的蛋白过表达。在本研究中发现的与茶足柄瘤蚜茧蜂滞育相关的蛋白质主要涉及烟酰胺腺嘌呤二核苷酸(NADH)脱氢酶亚基(复合物)、细胞色素bc1复合物亚基、ATP合酶ε亚基、谷氨酸脱氢酶(glutamate dehydrogenase, GDH)等。
NADH脱氢酶催化由NADH至辅酶Q的电子传递过程,同时将电子由线粒体基质转移至膜间隙。细胞色素bc1复合物是线粒体呼吸电子传递链中的核心元素,是催化从辅酶Q到细胞色素C的电子传递过程,同时将质子由线粒体基质泵至膜间隙。ATP合酶ε亚基,属于ATP合酶F1组分。ATP合酶,又称F0F1-ATP酶,在细胞内催化能源物质ATP的合成(王镜岩等, 2008)。在茶足柄瘤蚜茧蜂呼吸作用过程中通过电子传递链释放的能量先转换为跨膜质子(H+)梯差,之后质子流顺质子梯差通过ATP合酶可以使ADP+Pi合成ATP。ε亚基有抑制酶水解ATP的活性,同时有堵塞H+通道,减少H+外泄的功能,这一功能保证了茶足柄瘤蚜茧蜂在滞育过程中ATP的顺利合成(倪张林, 2001)。在环境胁迫条件下,能量需求增加,通过氧化磷酸化途径,能量产生增加,从而为滞育期间的茶足柄瘤蚜茧蜂提供更多的能量和营养物质,推断NADH脱氢酶、细胞色素bc1复合物、ATP合酶对茶足柄瘤蚜茧蜂的逆境生存和能量缓冲有积极作用。谷氨酸脱氢酶是调控机体碳、氮代谢相互交叉的重要酶,催化氧化脱氨基作用(oxidative deamination),氨基酸脱氨基后形成的氨是有毒物质。绝大多数陆生动物将脱下的氨转变为尿素排泄。在GO富集结果中显示,与天冬氨酸转运、L-谷氨酸转运条目相关的蛋白在滞育过程中上调表达,而天冬氨酸(aspartic acid)和谷氨酸(L-glutamic acid)是尿素形成的关键。线粒体中的谷氨酸脱氢酶将谷氨酸的氨基脱下,为氨甲酰磷酸(carbamoyl phosphate)的合成提供游离的氨;细胞质中的谷草转氨酶(aspartate aminotransferase)把谷氨酸的氨基转移给草酰乙酸(oxaloacetic acid),草酰乙酸再形成天冬氨酸进入尿素循环(urea cycle),谷氨酸为循环间接提供第2个氨基(王镜岩等, 2008)。同时谷氨酸脱氢酶在茶足柄瘤蚜茧蜂滞育蛹中上调表达,表明滞育蛹体内将有更多的氨进入尿素循环。这可能是由于滞育蛹新陈代谢较弱,从而抑制了氨基酸的合成,导致氨过剩的结果。此外,有研究报道,滞育型棉铃虫Helicoverpaarmigera幼虫可能通过在体内积累大量尿素达到抵御低温的作用(Zhangetal., 2013),茶足柄瘤蚜茧蜂滞育蛹也同样可能利用尿素来提高其耐寒性。NADH产生于糖酵解(glycolysis)和细胞呼吸作用中的柠檬酸循环(tricarboxylic acid cycle, TCA cycle),谷氨酸经过转化后可生成柠檬酸循环中间物质2-氧戊二酸,说明茶足柄瘤蚜茧蜂滞育对柠檬酸循环产生了显著影响。柠檬酸循环不仅为生命体提供能量,同时也是糖类、脂类和氨基酸三者之间互转化的枢纽(Bergetal., 2002)。已有研究表明,柠檬酸循环在昆虫滞育期间受到抑制(Michaud and Denlinger, 2007; Xuetal., 2012; 刘遥等, 2014; Luetal., 2014),这与滞育期间代谢减弱相符合。茶足柄瘤蚜茧蜂在滞育期间代谢减弱,与正常发育的茶足柄瘤蚜茧蜂相比,产能必定减少,但机体仍需要热能来抵御低温。在低温环境下,生物体产热增加,散热减少。而实验证明,在滞育过程中,参与氧化磷酸化通路的蛋白上调表达,说明此过程中产生的一部分能量用作维持正常的生命活动,还有一部分产生的是热能。自然界适应冷环境的动物,利用氧化磷酸化解偶联的方式产生大量的热。它们的脂肪组织中有一种褐色脂肪组织含有产热素(thermogenin)又称解偶联蛋白(uncoupling protein),能构建一种被动质子通道,使质子流从内膜外流向基质而不经过F0F1复合体的F0通道而是又回到基质,结果产生热而不形成ATP(王镜岩等, 2008)。我们推测,滞育的茶足柄瘤蚜茧蜂脂肪组织中可能也存在这种“解偶联剂”,使得虫体在氧化磷酸化过程中既能满足生命活动所需的能量,又能保证足够的热量来抵御低温环境。但滞育的茶足柄瘤蚜茧蜂体内是否含有这样的物质,我们需进一步探究。胆碱脱氢酶活性、胆碱生物合成甘氨酸甜菜碱条目相关蛋白在GO富集结果中显著上调,胆碱脱氢酶(choline dehydrogenase)可催化底物合成甘氨酸甜菜碱(glycine betain),因此甘氨酸甜菜碱的含量在滞育的茶足柄瘤蚜茧蜂蛹中必然增加。在滞育条件下,茶足柄瘤蚜茧蜂受到水分胁迫,甜菜碱作为有机渗透剂可维持细胞渗透压,同时甜菜碱对酶有保护作用,不仅可以抵御冰冻胁迫,对有氧呼吸和能量代谢过程也有良好的保护作用。
综上所述,本研究从蛋白质组整体层面阐明茶足柄瘤蚜茧蜂蛹滞育背后的多蛋白调控,重点筛选了与能量代谢相关的滞育关联蛋白并分析了其功能,有助于更好地理解茶足柄瘤蚜茧蜂蛹滞育的代谢机制,进一步扩展了对蚜茧蜂滞育机制的理解,为基于遗传或化学调控天敌滞育建立提供了新思路、新平台,具有重要的理论意义以及潜在的应用价值。
