春小麦抗旱耐热性QTL分析
2020-07-31白海波吕学莲马斯霜李树华董建力
白海波,吕学莲,惠 建,马斯霜,李树华,董建力
(宁夏农林科学院农业生物技术研究中心,宁夏银川 750002)
干旱和高温是制约小麦增产的两种主要非生物逆境,因此加强小麦抗旱耐热性遗传改良研究,对小麦高产稳产有重要意义。前人在小麦抗旱耐热性基因定位研究方面已经做了有意义的探索。张正斌等[1]利用小麦重组自交系(RILs)研究发现,控制小麦最大根长的QTL位于1B、2A、5A、5B、6A和7B上。周晓果等[2]以小麦DH群体在水分胁迫下检测到多个水分利用效率相关性状的QTL,它们分布在2A、3A、4A、5A、6A、7A、1B、3B、3D 染色体上。周瑞霞等[3]在干旱胁迫下检测到2个位于4B和7B染色体上、控制株高的QTL和2个均位于7B染色体上、分别控制穗茎节长和单株穗数的QTL。李卓坤等[4]以冬小麦DH群体为材料,在模拟水分胁迫条件下将控制小麦胚芽鞘长、根长的QTL定位在2A、4B和4D上。李世平等[5]利用冬小麦DH群体检测12个灌浆期耐热性QTL,其分布在1B、2D、3A、3B、6A、6B 和7A染色体上。Mason等[6]利用冬小麦RILs群体检测到小麦生育后期耐热性状的QTL位于1A、2A、2B、3B染色体上。Rajneesh等[7]利用RILs群体研究认为,小麦叶绿素含量、冠层温差、千粒重和产量的耐热指数的QTL分别位于2B、7B和7D上。Talukder等[8]发现,控制小麦叶绿素含量的QTL分别位于1B、1D、6A和7A染色体上。由于不同研究所用材料、性状、胁迫方式及环境不同,小麦抗旱、耐热QTL定位结果也不尽相同。
随着数量性状位点研究的深入,人们发现相关性状位点之间存在着大量的遗传重叠(genetic overlap),“一因多效”是导致性状间遗传重叠的一个重要原因。在双胁迫下水稻[9-11]、大豆[12]的抗旱与耐盐、耐盐与耐低温、抗病与抗旱等性状的QTL位点之间存在着大量的遗传重叠。本研究以抗旱、耐热小麦品种间杂交创制的RILs为作图群体,在干旱胁迫、热胁迫、旱热胁迫下对抗旱耐热相关性状QTL进行定位,确定QTL有利等位基因在染色体上的位置,并进行遗传重叠分析,以期为小麦抗旱耐热遗传改良及分子标记辅助选择(MAS)基因聚合育种提供依据。
1 材料与方法
1.1 试验材料
试验材料为“宁春4号×宁春27号”RILs群体(包括128个家系)及其亲本。宁春4号[13]由宁夏永宁育繁所选育,于1981年审定推广。宁春27号[14]由宁夏固原市农科所选育,于1998年审定推广。
1.2 试验设计
2015-2017年于宁夏农林科学院农作物所试验基地开展田间试验。每个材料种2行,行长1 m,行距0.2 m,顺序排列,重复3次。设干旱胁迫、热胁迫、旱热胁迫和对照(CK)4个处理。干旱胁迫处理仅在拔节期、抽穗期灌水,每次灌水量为900 m3·hm-2。干旱胁迫期间搭建遮雨棚架,雨前遮盖。热胁迫处理全生育期灌4水(拔节期、抽穗期、灌浆初期、灌浆中期),并在小麦灌浆中期38 ℃高温连续热胁迫处理3 d,每天3 h。增温方法采用李世平[5]、徐如强[15]的方法并略加改进,即用0.1 mm厚无色透明聚乙烯塑料人工搭建日光温棚,温棚顶部可移动调节棚内温度,在每天自然温度最高时段(12:00-15:00)进行热胁迫,在温棚穗层放置温度计、湿度计。旱热胁迫处理仅在拔节期、抽穗期灌水,灌浆初期至成熟干旱胁迫,灌浆中期进行热胁迫处理,方法与上面旱热单一胁迫相同。对照(CK)保持常温(自然温度),充足灌水,全生育期灌4水(拔节期、抽穗期、灌浆初期、灌浆中期),每次灌水量为900 m3·hm-2。
4个处理的施肥均一致,且与当地大田相同,每个处理间隔6 m,处理间在地下垂直埋塑料薄膜100 cm深,以防水分测渗,用量水堰监测灌水量。灌水处理的小区搭建遮雨棚,在雨前人工遮盖。
1.3 测定项目及方法
1.3.1 叶绿素含量测定
在干旱、热胁迫前后用SPAD-502 叶绿素仪于上午9: 00-12:00在各小区第一行测同一天抽穗挂牌的5个主茎的旗叶上、中、下部SPAD值,取其平均值。
1.3.2 叶片含水量测定
在干旱、热胁迫前后取各小区第一行的主茎旗叶5片,随即称鲜重,置于70 ℃烘箱烘干,称干重,计算旗叶含水量(RWC)。RWC=(鲜重-干重)/干重×100%。
1.3.3 穗粒重、千粒重测定
成熟期,在每小区第二行收10个主穗,考察穗粒重、千粒重。
1.4 遗传连锁图谱的构建及QTL定位分析
用分布于小麦21对染色体上的307对多态性SSR标记对RILs群体128个株系进行基因型分析,利用完备区间作图软件IciMapping 4.0[16],构建包含30个连锁群的遗传图谱(图1)。图谱包括266个标记位点,遗传连锁图总长度为 2 187.79 cM,平均每条染色体连锁图长104.18 cM,平均2个标记的距离为8.22 cM[17]。

图1 小麦SSR遗传连锁图谱
采用IciMapping 4.0对RILs群体抗旱耐热相关性状进行QTL定位,以LOD>3.0作为阈值判断QTL存在与否,以α=0.05和α=0.01为显著水平计算表型贡献率。加性效应为正值,表明QTL的增加效应来自于母本宁春4号;加性效应为负值,表明其增加效应来自于父本宁春27号。测定数据采用 Microsoft Excel 2007软件进行统计分析。
2 结果与分析
2.1 RILs及其亲本性状表现
3种胁迫条件下,亲本和RILs群体间旗叶叶绿素含量、旗叶含水量、穗粒重和千粒重的差异较大,各性状变化均存在超亲现象,各性状值的偏度和峰度绝对值小于1或接近1,w值接近1(表1),表明RILs群体的生理性状、产量性状值呈正态分布,属于数量性状遗传,适合进行QTL 定位。
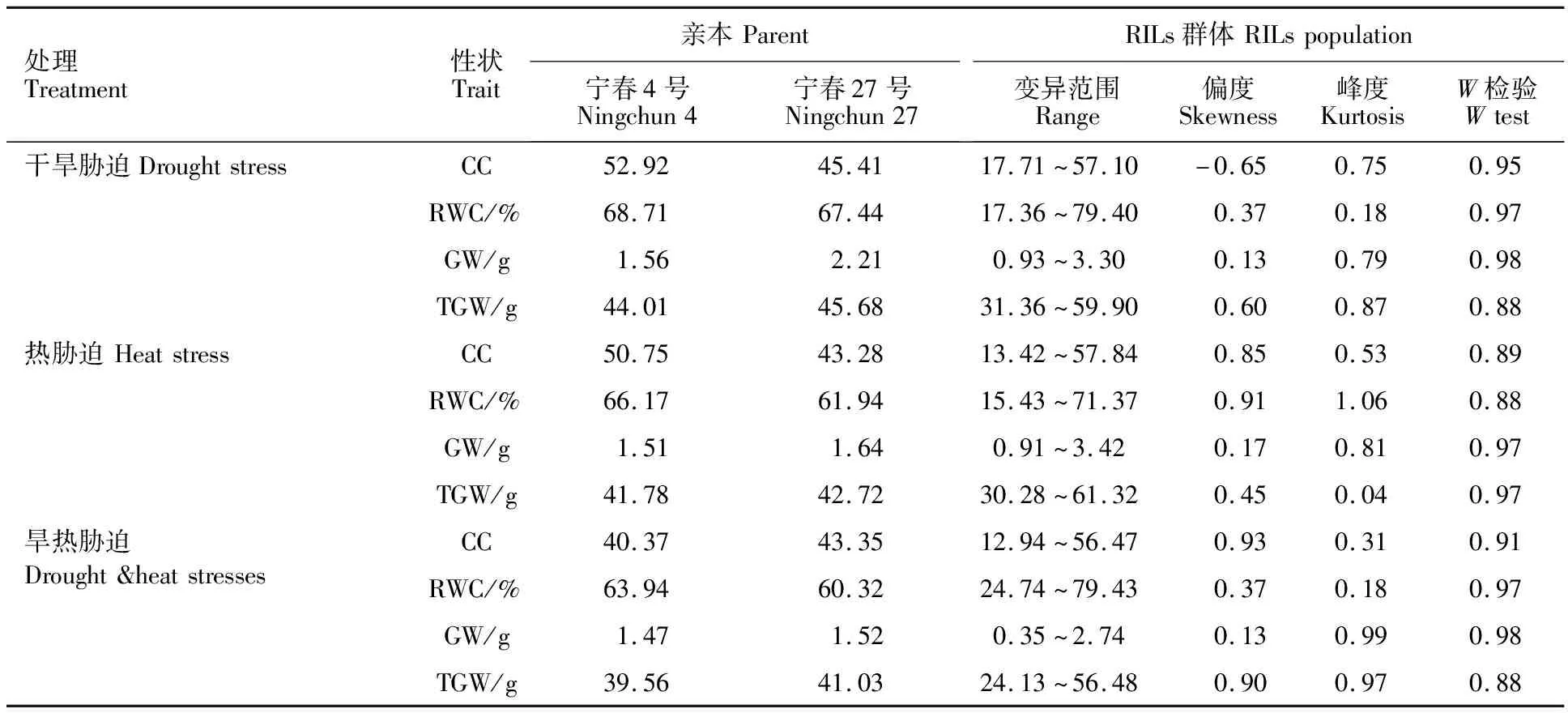
表1 不同胁迫下亲本和RILs群体性状表现
2.2 灌浆期抗旱相关性状的 QTL定位结果
在三年灌浆期干旱胁迫下,共检测到控制旗叶叶绿素含量、旗叶含水量、千粒重和穗粒重的QTL 22个,它们分布于2A、3B、4B、7B、3D、4D、7D染色体上,单个QTL的表型贡献率为 9.38%~30.81%(表2)。控制旗叶叶绿素含量的QTL有5个,位于2A、3B、4B和7B染色体上,表型贡献率为 12.45%~21.46%;其中Qcc-2A.2和Qcc-7B等位基因来自抗旱父本宁春27号,表型贡献率为11.93%~21.46%。控制旗叶含水量的QTL有7个,位于2A、3B、3D、4D、7B和7D染色体上,表型贡献率为9.38%~30.81%;其中Qrwc-2A、Qrwc-3B.1、Qrwc-3D和Qrwc-7D等位基因来自抗旱父本宁春27号,表型贡献率为9.38%~ 30.54%。控制穗粒重的QTL有6个,位于2A、3B、4B、4D 和7B染色体上,表型贡献率为9.76%~25.42%;其中Qgw-3B、Qgw-4B.2、Qgw-4D和Qgw-7B等位基因来自抗旱父本宁春27号,表型贡献率为9.76%~25.42%。控制千粒重的QTL有4个,位于2A、7B、3D和7B染色体上,表型贡献率为9.75%~25.83%;其中Qtgw-2A和Qtgw-3D等位基因来自抗旱父本宁春27号,表型贡献率分别为9.75%和13.18%。2A和7B染色体上各有1个QTL在三年干旱胁迫处理中均被检测到,分别控制旗叶含水量(Qrwc-7B,wmc83-wmc276)和穗粒重(Qgw-2A,gwm294-wmc644)。有6个QTL在两年被检测到,其中2个控制旗叶叶绿素含量(Qcc-2A.2、Qcc-4B),1个控制旗叶含水量(Qrwc-4D),2个控制穗粒重(Qgw-3B、Qgw-4B.2),1个控制千粒重(Qtgw-2B)。

表2 干旱胁迫下抗旱相关性状QTL定位
2.3 灌浆期耐热相关性状的QTL定位结果
3年热胁迫下,共检测到控制旗叶叶绿素含量、旗叶含水量、千粒重和穗粒重的QTL 36个,它们分布于2A、2B、3A、3B、4B、4D、5D、6D、7A和7B染色体上,单个QTL的表型贡献率为 9.03%~34.97%(表3)。控制旗叶叶绿素含量的QTL有12个,位于2A、3A、3B、4B、5D、6D、7A和7B染色体上,单个QTL的表型贡献率为 9.39%~34.16%,除了Qcc-3B外,其他等位基因来自耐热亲本宁春4号。控制旗叶含水量的QTL有8个,位于2A、2B、4D、6D、7A和7B染色体上,单个QTL的表型贡献率为9.42%~ 43.73%,等位基因全部来自耐热亲本宁春4号。控制穗粒重的QTL有8个,位于2A、3B、4D、5D、6D和7A染色体上,单个QTL的表型贡献率为9.03%~24.29%,2A、4D、5D、6D和7A染色体上等位基因来自宁春4号,3B染色体上等位基因来自父本宁春27号。控制千粒重的QTL有8个,位于2A、2B、3B、4B、5D和6D染色体上,单个QTL的表型贡献率为9.49%~42.57%,4B、5D和6D染色体上等位基因来自宁春4号。3B、7B染色体上各有1个QTL在三年热胁迫处理中均被检测到,分别控制穗粒重(Qgw-3B)和旗叶含水量(Qrwc-7B.1)。

表3 热胁迫下耐热相关性状QTL定位
2.4 灌浆期旱热胁迫下QTL定位结果
灌浆中期旱热胁迫下,共检测到控制旗叶叶绿素含量、旗叶含水量、千粒重和穗粒重的QTL 30个,它们分布于2A、3A、4A、2B、3B、4B、7B、3D、4D、6D染色体上,单个QTL的表型贡献率为9.09%~40.81%(表4)。控制旗叶叶绿素含量的QTL有9个,位于2A、2B、3A、3B、4A和4D染色体上,单个QTL的表型贡献率为9.10%~33.01%;其中,Qcc-2A.1、Qcc-2A.2、Qcc-2B.2、Qcc-3A和Qcc-4D等位基因来自宁春4号,Qcc-2B.1、Qcc-3B、Qcc-3D、Qcc-4D等位基因来自宁春27号。控制旗叶含水量的QTL有6个,位于2B、3B、4B、7B染色体上,单个QTL的表型贡献率为13.92%~55.68%,等位基因全部来自耐热亲本宁春4号。控制穗粒重的QTL有8个,位于2A、3B、4B、6D和7B染色体上,单个QTL的表型贡献率为8.40%~35.67%;其中,Qgw-2A.1、Qgw-2A.2、Qgw-4B.1和Qgw-6D等位基因来自宁春4号,Qgw-2B、Qgw-3B.1、Qgw-3B.2、Qgw-4B.2和Qgw-7B等位基因来自宁春27号。控制千粒重的QTL有7个,位于2B、3D、4D、6D 和7B染色体上,单个QTL的表型贡献率 为8.81%~39.01%;其中,Qtgw-6D.1和Qtgw-6D.2等位基因来自父本宁春4号。2A、3B、7B染色体上各有1个QTL在三年旱热胁迫处理中被检测到,分别控制旗叶叶绿素含量(Qcc-2A.2)、旗叶含水量(Qrwc-7B.1)和千粒重(Qtgw-2B)。

表4 旱热胁迫下QTL定位
2.5 抗旱耐热性共定位QTL
2.5.1 抗旱相关性状共定位QTL
在小麦灌浆期干旱胁迫下,2A染色体上检测到的控制旗叶叶绿素含量的QTLQcc-2A.2与穗粒重的QTLQgw-2A共定位区间为 gwm294-wmc644,区间距为6.49 cM(图2),表型贡献率为13.13%~ 21.46%;7B染色体上检测到的控制旗叶叶绿素含量的QTLQcc-7B与穗粒重的QTLQgw-7B共定位区间为barc140-gwm297,区间距为1.92 cM,表型贡献率为11.03%~ 12.45%。这两个共定位区间控制性状的加性效应均为负值,其增效等位基因来自抗旱亲本宁春27号,说明该QTL位点与灌浆期抗旱性的关系密切,其中共定位区间gwm294-wmc644在2年均被检测到,表明该位点受环境的影响较小,能够稳定表达。
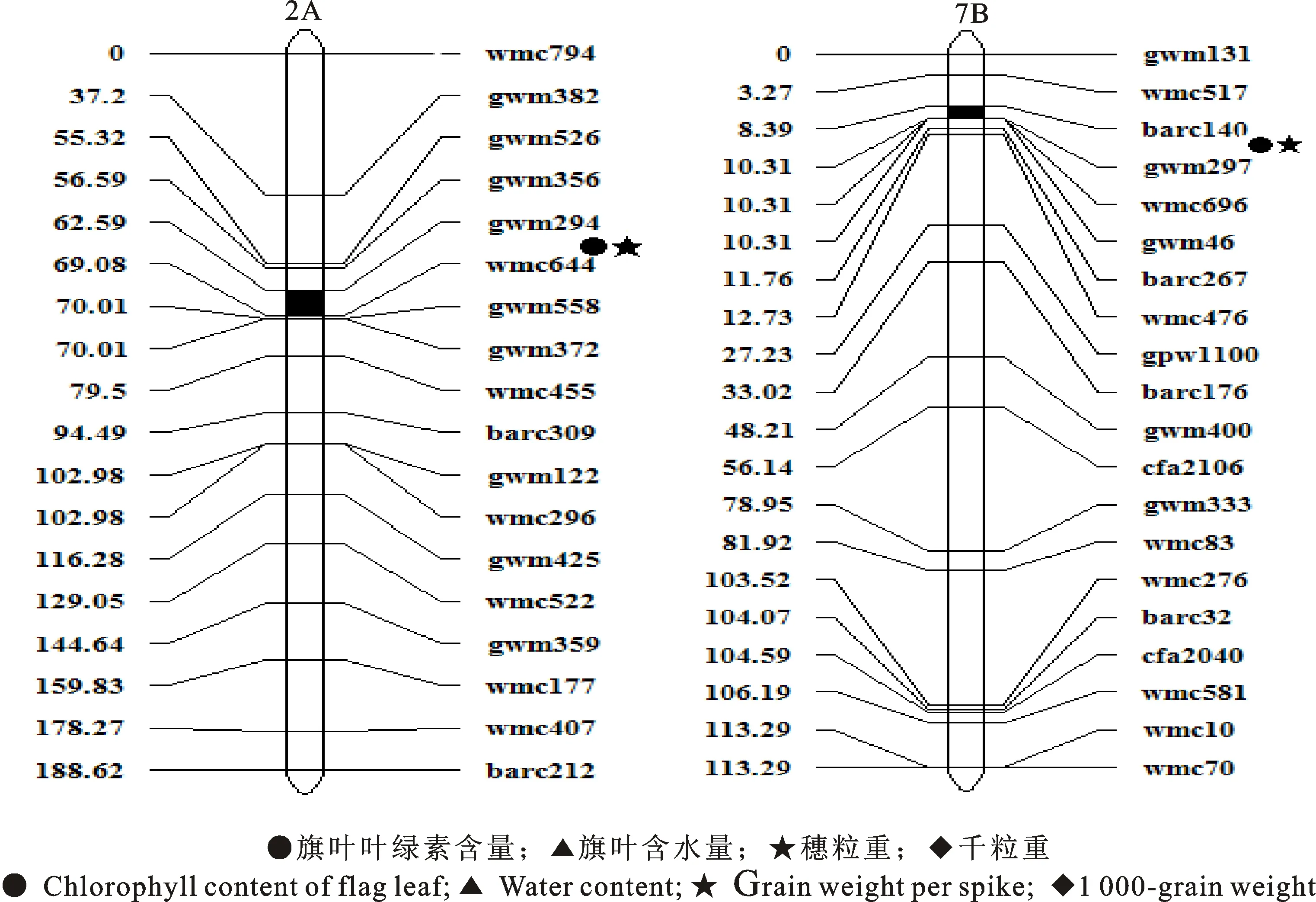
图2 小麦抗旱性QTL共定位区间在染色体上的分布
2.5.2 耐热相关性状共定位QTL
在小麦灌浆期热胁迫条件下,检测到5D染色体上控制旗叶叶绿素含量的QTLQcc-5D与穗粒重的QTLQgw-5D.1的共定位区间为cfd67-cfd40(图3),区间距为11.24 cM,表型贡献率为16.57%~18.28%。在6D染色体上检测到的控制旗叶含水量的QTLQrwc-6D、穗粒重的QTLQgw-6D.1和千粒重的QTLQtgw-6D.1共定位区间为barc196-barc54,区间距为4.71 cM,表型贡献率为9.42%~16.59%。加性效应分析表明,共定位区间 cfd67-cfd40和 barc196-barc54控制性状的加性效应均为正值,其增效等位基因来自耐热亲本宁春4号,说明其与灌浆期耐热性关系密切,但因为只在一年一种胁迫下检测到,因而其表达受环境的影响较大。

图3 小麦耐热性QTL共定位区间在染色体上的分布
2.5.3 抗旱性QTL与耐热性QTL共定位分析
在小麦灌浆期旱热胁迫下,检测到2A染色体上控制旗叶叶绿素含量的QTLQcc-2A.2与穗粒重的QTLQgw-2A.1共定位区间为gwm294-wmc644(图4),表型贡献率为10.13%~ 26.14%。加性效应分析表明,在干旱胁迫下控制旗叶叶绿素含量的等位基因来自抗旱亲本宁春27号,在热胁迫下控制穗粒重的等位基因来自耐热亲本宁春4号,说明QTL位点 gwm294-wmc644与抗旱耐热性有密切关系,且在干旱胁迫、热胁迫、旱热胁迫条件下都被检测到,说明该位点受环境的影响小。
在2B染色体上检测到的控制旗叶叶绿素含量的QTLQcc-2B与千粒重的QTLQtgw-2B共定位区间为wmc441-wmc317(图4), 表型贡献率为13.27%~ 22.13%。加性效应分析表明,控制千粒重的等位基因来自宁春27号,控制旗叶叶绿素含量的等位基因来自宁春4号,说明该位点与抗旱耐热性有密切关系。
在7B染色体上,控制旗叶含水量的QTLQrwc-7B.1与千粒重的QTLQtgw-7B的共定位区间为wmc83-wmc276(图4),表型贡献率为 27.21%~55.68%,遗传效应较大。经加性效应分析,控制旗叶含水量的等位基因来自宁春4号,控制千粒重的等位基因来自宁春27号,共定位区间 wmc83-wmc276与抗旱耐热性有密切关系,且在2年三种胁迫下被检测到,说明该位点受环境的影响小。

图4 小麦抗旱耐热性QTL共定位区间在染色体上的分布
3 讨 论
小麦的抗旱耐热性是由多基因控制的复杂数量性状,是小麦本身的遗传特性和环境共同作用的结果。本研究与前人在2A[1,2,4]、7B[1-3,18]、4B[4-5]染色体上检测到与抗旱性有关的QTL结果相同,与赵 朋等[19]和刘胜男等[20]都在2A染色体上检测到控制叶绿素含量的QTL结果一致,本研究所得的叶绿素含量QTLQcc-2A.2的染色体区间(gwm294-wmc644)也与刘胜男等定位的叶绿素含量Qcc.ahau2-as2[20-21]、赵 朋等[19]定位的叶绿素含量Qcc-2A染色体区间相吻合。
在耐热基因定位方面,本研究与Mason等[6]在2A、3B染色体上及与李世平等[5]、Pinto等[22]在7A染色体上检测到与耐热性有关的QTL结果相同,但与Rajneesh等[7]和Vijayalakshmi等[23]在7B染色体上检测到与耐热性有关的QTL结果不一致。这种差异可能与试验材料、鉴定方法以及QTL定位工具等的不同有关。
目前,关于小麦叶绿素含量、穗粒重、千粒重QTL定位的研究报道较多,而对小麦组织含水量QTL定位的研究报道极少。本研究发现,小麦灌浆期热胁迫下控制旗叶含水量、千粒重的QTL均定位在6D染色体 barc196-barc54区间内,均未与前人研究报道的位点重合或相近。这可能与小麦的基因组较大、抗逆性遗传比较复杂有关,因此应该开展多年多胁迫下小麦抗逆性QTL定位,以发掘更多的抗逆遗传信息和功能分子标记。
近年来,关于小麦抗旱、耐热相关性状QTL定位研究报道较多,但大都局限于单一胁迫。本研究中,小麦抗旱性共定位区间有2个,分别位于2A(gwm294-wmc644)和7B(barc140-gwm297)染色体上;耐热性共定位区间有2个,分别位于5D(cfd67-cfd40)和6D(barc196-barc54)染色体上;抗旱耐热性共定位区间有3个,位于2A(gwm294-wmc644)、2B(wmc441-wmc317)和7B(wmc83-wmc276)染色体上。小麦抗旱耐热性共定位位点 gwm294-wmc644(Qcc-2A.2、Qrwc-2A.1)的贡献率大、遗传距离较小(6.49 cM)。抗旱耐热性QTL定位发现,在干旱胁迫、热胁迫、旱热胁迫下控制旗叶含水量QTL的平均贡献率位居第一,叶绿素含量的平均贡献率位居第二,表明旗叶含水量和叶绿素含量对小麦抗旱耐热具有重要作用。
