普通小麦-滨麦及其衍生系的染色体组成分析
2020-07-31杨晓菲王长有陈春环田增荣吉万全
杨晓菲,王长有,陈春环,田增荣,吉万全
(1.渭南师范学院环境与生命科学学院,陕西渭南 714099; 2.旱区作物逆境生物学国家重点实验室/西北农林科技大学农学院,陕西杨凌 712100)
赖草属(LeymusHochst.)是一个从四倍体至十二倍体类型均有的多倍体物种,其中许多物种[如大赖草(L.racemusus)、羊草(L.chinensis)、多枝赖草(L.multicaulis)等]的染色体核型已经非常清楚[1-2],而滨麦(Leymusmollis)是赖草属中的一个四倍体物种,具有抗寒、抗旱、耐盐碱[3]、茎秆粗壮和大穗[4]等优良性状,同时对小麦条锈病、叶锈病和白粉病等多种真菌病害免疫[5-6]。作为小麦改良育种的重要三级基因资源[7],其染色体核型的研究处于空白状态,且四倍体赖草属植物基因组(染色体组)构成目前尚未定论。滨麦基因组为NsNsXmXm,其中Ns基因组来自于新麦草(PsathyrostachysKeng)[8-10],而Xm基因组的来源目前尚未确定。在第2届国际小麦族研讨会上(1994年美国犹他州),基因组命名委员会建议使用NsXm作为四倍体赖草属植物基因组的表示符号,Xm代表一个未知的基因组直到试验验证为止[11]。
染色体组型分析可以初步判断外源染色体是否存在和具体数目,是植物分类和遗传研究的重要方法之一。本研究对滨麦染色体核型进行初步分析并绘制滨麦染色体核型模式图,同时通过分子标记研究其可能的二倍体供体种并分析其与滨麦之间的亲缘关系。
Pedersen等[12]利用FISH技术结合重复序列pAsl和pHvG38,分辨出中国春和绵阳11中的21对染色体。由于重复序列探针的准备耗时费力,部分寡核苷酸探针的发展为辨别染色体提供了一种更经济高效的方法。Tang等[13]研究认为,两种寡核苷酸探针pSc119.2和pTa-535代替重复序列pAsl和pHvG38做双色FISH,同样可以清晰准确地识别小麦21对染色体。因此,本研究拟结合两种寡核苷酸探针对普通小麦7182的标准核型进行绘制,并利用FISH-GISH技术对普通小麦-滨麦高世代衍生系的染色体组成进行分析,旨在为后续创制和鉴定出更多类型的普通小麦-滨麦种质资源奠定基础。
1 材料与方法
1.1 试验材料
研究材料包括普通小麦7182(Triticumaestivum,2n=6x=42,AABBDD),滨麦(L.mollis,2n=4x=28,NsNsXmXm),华山新麦草(Psathyrostachyshuashanica,2n=2x=14,NsNs),二倍体长穗偃麦草(Thinopyrumelongatum,2n=2x=14,EeEe),拟鹅观草(Pseudoroegeriaspicata,2n=2x=14,StSt)和百萨偃麦草(Thinopyrumbessarabicum,2n=2x=14,EbEb)以及普通小麦-滨麦高世代衍生系M47和M39。
1.2 试验方法
1.2.1 根尖培养与取材
将供试材料种子腹沟朝下,置于铺有两层滤纸的干净培养皿中,加ddH2O湿润滤纸即可(无多余的水分流动为宜)。将培养皿置于23~25 ℃恒温培养箱中黑暗培养,待根长至3~5 cm时,剪取长约1 cm根尖放入湿润且打孔的普通离心管(1.5 mL)中并编号记录,放入笑气(N2O)罐中密封处理2 h,之后在离心管中加入300 μL 90% 冰乙酸,冰浴固定10 min后,ddH2O冲洗根尖3次,静置,待离心管完全干燥后,加入70% 冰乙醇,置于-20 ℃冰箱保存备用。
滨麦根尖于2015-2017连续三年在3月下旬至4月上旬取材两到三次,挖取新生根并剪取约1 cm长的根尖,置于冰水混合物中处理22~26 h,然后用卡诺氏固定液(无水乙醇∶冰乙酸=3∶1,v/v)于4 ℃冰箱固定48 h左右,用ddH2O将根尖冲洗至少3次,随后加入70%冰乙醇,-20 ℃冰箱保存备用。
1.2.2 根尖处理与制片
用ddH2O冲洗70%冰乙醇保存的根尖,用滤纸吸干水分,切取乳白色根尖部分置于装有2% 纤维素酶和1%果胶酶混合酶液的0.5 mL离心管中,瞬时离心,待乳白色根尖沉入离心管底部,将离心管放入37 ℃水浴酶解55~60 min(不同材料酶解时间有所不同)。然后用70% 乙醇冲洗三次,用解剖针将浸润在100 μL 70% 乙醇中的根尖捣碎,6 000 r·min-1离心1~2 min,倒置离心管于滤纸上吸干水分,加40 μL预冷的纯乙酸,高速瞬时涡旋3次,移液枪吸取10 μL悬浊液滴于干净载玻片上(处于黑暗且湿润的盒子),5 min后待悬浊液缓慢均匀扩散后于显微镜下镜检并拍照。
1.2.3 染色体核型分析与模式图绘制
根据李懋学等[14]关于植物核型的分析标准,选取30个以上染色体形态和分散较好的中期分裂相细胞进行染色体数目的统计,精选五个细胞进行拍照。利用图象处理软件Adobe Photoshop CS6进行细胞背景处理,测量各个染色体及各臂的相对长度,进行染色体的配对,排列建立核型图。
利用Excel对染色体的各种参数进行数据分析,并绘制出整套染色体的核型模式图。按照Stebbins[15]的核型标准进行核型分类。
1.2.4 SSR和EST分子标记分析
分子标记分析共选用825对SSR引物和86对EST引物,SSR和EST引物序列均来自GrainGenes数据库(http://wheat.pw.usda.gov/GG2/index.shtml),由奥科鼎盛生物科技有限公司(北京)合成。采用改良后的CTAB法对供试材料进行总基因组DNA的提取。PCR反应体系为10 μL,包括1.0 μL PCR Buffer 缓冲液(含Mg2+)、1.0 μL 0.5 μmol·L-1primers(正、反向引物各0.5 μL)、0.8 μL 0.2 mmol·L-1dNTPs混合液、模板DNA 1.0 μL(200 ng·μL-1)、0.1 μL Taq DNA 聚合酶(5 U·μL-1),ddH2O补充至 10.0 μL。反应在热循环仪Bio-Rad S1000TM型(California,USA)上进行。PCR产物用8%非变性聚丙烯酰胺凝胶检测,电泳步骤参照吴金华等[16]的方法,凝胶经硝酸银染色显影后置于灯箱上观察、照相并统计带型。
1.2.5 双色FISH-GISH技术
参照Tang 等[13]的方法,利用英骏(Invitrogen)生物技术有限公司(上海)合成的两种寡核苷酸探针Oligo-pTa535(Tamra-red)、Oligo-pSc119.2(FAM-green)序列,以及滨麦叶片总基因组DNA为探针,先后对根尖体细胞进行荧光原位杂交(FISH)和基因组原位杂交(GISH)分析。滨麦叶片gDNA的提取参照改良后的CTAB法[17]进行。使用OLYMPUS BX53(日本)荧光显微镜镜检,利用系统软件DP800 CCD、cellSens Standaed 1.8进行拍照并用图像处理软件Adobe Photoshop CS6分析图像。
2 结果与分析
2.1 滨麦染色体核型分析及核型模式图
镜检结果表明,滨麦根尖的细胞染色体数目为28条,即2n=4x=28(图1a)。利用图像处理软件Adobe Photoshop CS6对染色体及各臂的相对长度进行测量,根据测量结果进行配对和排序,绘制滨麦染色体核型图(图1b)。计算染色体长臂与短臂相对长度的比值并确定每一对染色体所属类型(表1),绘制出滨麦染色体核型模式图(图1c)。依据核型标准推测出滨麦的核型公式为:2n=4x=28=22m(6sat)+6sm,其中m、sm和sat分别代表中间着丝粒染色体、亚中间着丝粒染色体和具有随体的染色体。
2.2 滨麦染色体核型的归属
根据滨麦各对染色体的臂比(表1,图1c),发现中间着丝粒染色体和亚中间着丝粒染色体分别有11对和3对。依据Stebbins[15]的染色体核型标准分类方案,即最长与最短染色体之间的长度比值<2记为A级,比值在2~4之间记为B级,比值>4记为C级;臂比(长臂/短臂)>2的比例为0记为I级,0.01~0.5记为II级,0.51~0.99记为Ⅲ级,1.0记为Ⅳ级。本研究中滨麦最长和最短染色体的比值为1.98,记为A级;染色体臂比>2的比例为0.07,记为II级。因此,滨麦染色体的核型类型为IIA。
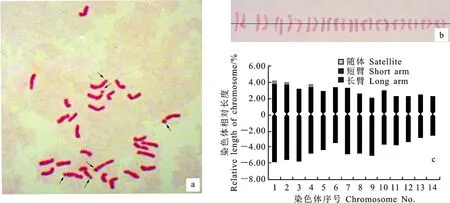
箭头指示滨麦染色体的随体。

表1 滨麦染色体的相对长度(总长、长短臂)、臂比及类型
2.3 滨麦染色体组供体种的初步分析
分子标记扩增结果表明,456对标记(包括379对SSR引物和77对EST引物)在二倍体华山新麦草、二倍体长穗偃麦草、百萨偃麦草、拟鹅观草和四倍体滨麦中表现多态性(部分结果如图2),58对标记在滨麦和四种二倍体物种中扩增带型完全一致,其余311对标记在滨麦、华山新麦草、二倍体长穗偃麦草、拟鹅观草和百萨偃麦草中无扩增条带或者扩增条带模糊。滨麦染色体组供体种分子标记的初步分析结果表明,在多态性标记中有157对SSR标记和27对EST标记在华山新麦草和滨麦间扩增出相同带型;有52对SSR标记和15对EST标记在百萨偃麦草和滨麦间扩增出相同带型;有62对SSR标记和21对EST标记在二倍体长穗偃麦草和滨麦间扩增出相同带型;有67对SSR标记和24对EST标记在拟鹅观草和滨麦间扩增出相同带型。这说明华山新麦草、百萨偃麦草、二倍体长穗偃麦草和拟鹅观草与滨麦扩增出相同带型的标记分别占多态性标记数的40.35%、14.69%、18.20% 和19.96%。

a:Xgpw7574-5B标记的PCR结果;b:Xgwm674-3A标记的PCR结果;M:Marker;1:滨麦;2:华山新麦草;3:百萨偃麦草;4:二倍体长穗偃麦草;5:拟鹅观草。
2.4 普通小麦7182的FISH标准核型图
傅 杰等[18]通过组织培养获得普通小麦-滨麦的杂种F1植株,经秋水仙素处理后与普通小麦品系7182回交获得了杂种后代。本研究结合两种寡核苷酸探针pSc119.2和pTa-535,采用双色FISH技术绘制出普通小麦7182所有染色体的FISH标准核型图(图3)。

红色:Oligo-pTa535探针;绿色:Oligo-pSc119.2探针;蓝色:DPAI复染。
2.5 普通小麦-滨麦高世代衍生系M47和M39的染色体组成
利用寡核苷酸探针pSc119.2和pTa-535对高世代衍生系M47和M39进行顺序双色FISH-GISH鉴定。细胞学观察结果表明,M47和M39体细胞染色体数目均为56条,不同的是,M47中有42条普通小麦染色体,14条双末端红色信号染色体(图4a);而M39中有44条普通小麦染色体,其中包含4条4B小麦染色体,仅有12条双末端红色信号染色体(图5a),因此,初步推断双末端红色信号染色体为滨麦染色体。为了进一步确认双末端红色染色体的来源,利用滨麦的总基因组DNA为探针进行GISH鉴定,结果表明,双末端红色信号染色体为滨麦染色体(图4b、5b)。
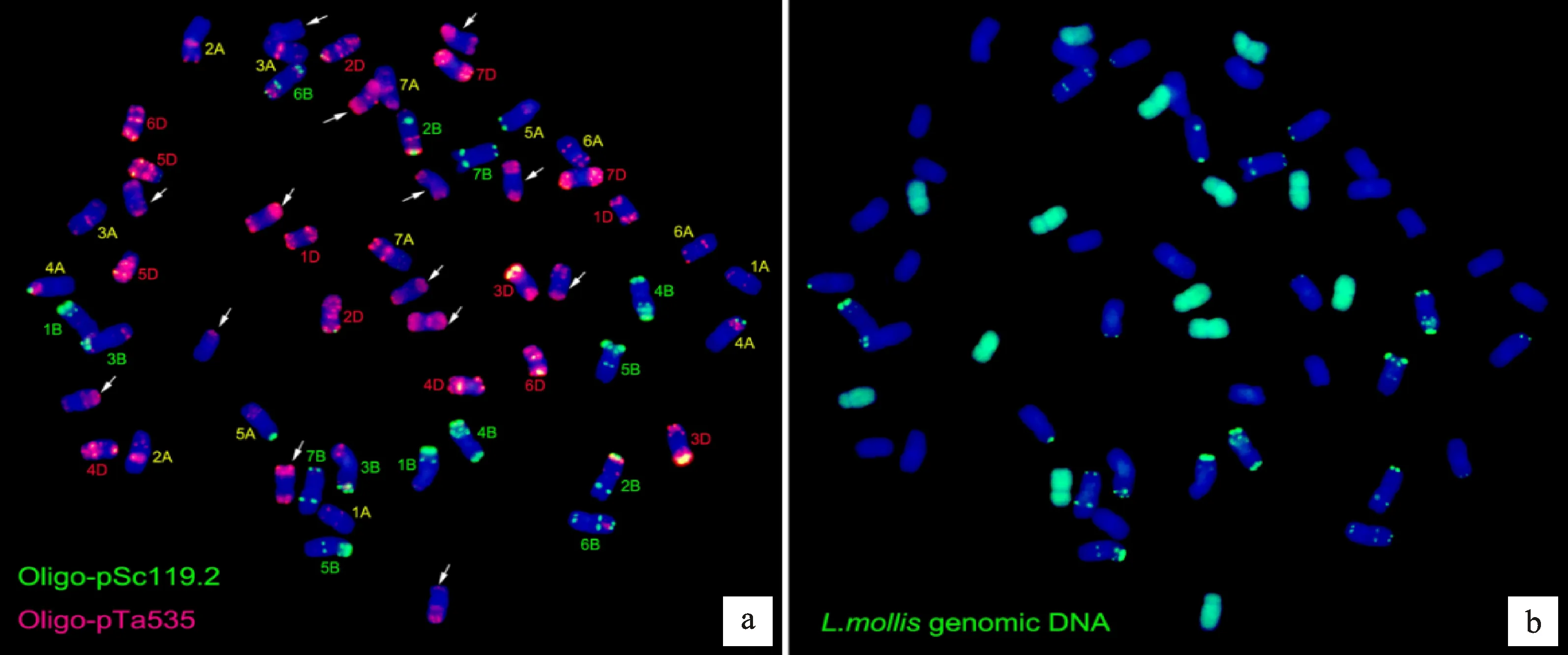
a:利用寡核苷酸探针Oligo-pTa535(红色)和Oligo-pSc119.2(绿色)对M47的FISH分析,显示42条小麦染色体(已标出)及14条双末端红色信号染色体(白色箭头所指);b:利用滨麦总基因组DNA 为探针(绿色)在同一张片子上对M47的GISH分析。以DAPI为复染剂(蓝色)。

a:利用寡核苷酸探针Oligo-pTa535(红色)和Oligo-pSc119.2(绿色)对M39的FISH分析,显示44条普通小麦染色体(已标出)包含4条4B染色体(白色大箭头所指)及12条双末端红色信号染色体(白色小箭头所指);b:利用滨麦总基因组DNA 为探针(绿色)在同一张片子上对M39的GISH分析。DAPI为复染剂(蓝色)。
3 讨 论
3.1 滨麦核型及染色体组构成分析
智 力[19]于2000年报道了窄颖赖草(L.angustus)、粗穗赖草(L.crassiuculus)、若羌赖草(L.ruoqiangensis)、大赖草(L.ruoqiangensis)和羊草(L.chinensis)等5个不同赖草属物种的染色体核型;杨瑞武等[1]于2004年报道了十一个赖草属四倍体物种,包括多枝赖草(L.multicaulis)、分枝赖草(L.ramosus)、沙生赖草(L.arenarius)、盐生赖草(L.salinus)、密穗赖草(L.condensatus)、新生赖草(L.innovatus)、灰赖草(L.cinereus)、柴达木赖草(L.pseudoracemosus)、无芒赖草(L.triticoides)、阿克摩林赖草(L.akmolinensis)及赖草(L.secalinus)的染色体核型。分析比较得知,上述赖草属不同物种均具有随体染色体,且多数为中部着丝粒染色体,每个物种具有数量不等的近中部着丝粒染色体,其中个别物种还有少数的近端部着丝粒染色体。从核型的分类类型来看,除多枝赖草和沙生赖草核型为IIB外,其余物种的核型均为IIA。而葛荣朝等[20]发现,多枝赖草染色体核型中未出现随体染色体,这与杨瑞武[1]的研究结果不一致,造成结果差异的原因可能与试验材料来源和所采用的试验方法不同以及试验操作过程中的误差等有关。
本研究对四倍体滨麦进行了核型分析,核型公式为2n=4x=28=22m(6sat)+6sm,核型类型为ⅡA,可以得出具有随体的滨麦染色体以中部着丝粒染色体为主,有少数的近中部着丝粒染色体,这与前人报道的赖草属大多数物种的核型相似。综合分析,赖草属不同物种的染色体核型既有相似之处,也有各自的特征差异,这种差异可以作为物种基础分类的细胞学依据。多态性分子标记在华山新麦草、百萨偃麦草、二倍体长穗偃麦草和拟鹅观草与滨麦扩增出相同带型的不同比例初步表明,华山新麦草与滨麦的亲缘关系较其他三个供体种更近。
3.2 普通小麦7128标准核型及其与滨麦高世代衍生系的染色体组成
本研究利用两种寡核苷酸探针pSc119.2和pTa-535代替耗时费力的重复序列pAsl和pHvG38,更加经济、高效和准确地绘制出普通小麦7182中21对染色体的标准FISH核型图。通过双色FISH-GISH技术鉴定出两种类型的普通小麦-滨麦高世代衍生系M47(2n=56=42T.a+14L.m)和M39(2n=56=44T.a+12L.m),均可作为创制新的普通小麦-滨麦种质资源的中间桥梁材料。本研究为后续普通小麦-滨麦种质资源的创制和鉴定提供了基础理论依据和技术 方法。
