糜子矮秆突变体“819”矮秆基因的遗传学分析
2020-03-07贾小平王振山朱学海杨德智寇淑君刘星星
贾小平,王振山,朱学海,杨德智,寇淑君,刘星星
(1.河南科技大学 农学院,河南 洛阳 471000; 2.张家口市农业科学院 黍子研究所,河北 张家口 075000)
由于矮秆材料具有抗倒伏和可实现密植的优点,发掘新的矮秆资源成为作物育种改良和高产作物研究的重要工作。农业生产上出现的第一次“绿色革命”便是利用了矮化的小麦植株与墨西哥抗锈病小麦杂交,最后培育出具有抗倒伏高产特性的小麦新品种,大幅度提高了世界粮食的产量,不仅暂时解决了粮食短缺的问题,还带来了高新技术的发展和丰厚的经济利益[1]。通过对玉米的大量研究表明,赤霉素生物合成代谢的通路与植株的高度有关,依据对外源赤霉素反应的差异程度,可以将矮秆突变体分为2种类型:赤霉素钝感型和赤霉素敏感型,由于赤霉素生物合成途径酶的缺陷所导致的矮秆突变称为赤霉素敏感型矮秆突变,对外源赤霉素的施加反应不明显的突变体叫赤霉素钝感型矮秆突变[2]。水稻的有关研究结果表明,籼稻的矮秆性状主要受1个隐性半矮秆基因所控制[3]。粳稻中矮秆基因控制的矮秆性状可分为2种,一种是由1个矮秆主效基因所控制,一种是由多个矮秆微效基因所控制[4-5]。整体来看,水稻中已经发现的矮秆基因中隐性基因遗传的占绝大部分,只有极少数呈部分显性或显性遗传[6]。小麦在我国的种植面积仅次于水稻[7],目前已发掘出来的小麦矮秆基因主要分为3种:第1种是丛簇矮生性基因(D),第2种是单秆矮生基因(Us),这2种矮生资源在育种实践中应用不多,而第3种主效降低株高基因(Rht)是生产上应用最普遍的基因资源[8]。
糜子是一种古老的C4作物,生育期为60~100 d,由于具有强的耐旱性和高水分利用效率,适合在我国北方干旱和半干旱地区种植。糜子籽粒又称为黄米,富含蛋白质、维生素和矿质元素,具有较高的营养价值,加工的产品称油糕或年糕,在我国山西、内蒙、甘肃、河北等北方地区深受广大人民群众的欢迎,特别是过年、婚庆等喜庆节日,都会作为一种必不可少的食物接待宾客,消费市场广阔。然而由于糜子抗倒伏能力较弱,在多风、干旱丘陵地区极易发生大面积倒伏,造成严重减产,创新矮秆资源、发现矮秆基因、培育矮化糜子新品种是解决这一问题的根本途径。近些年有关糜子分子标记的研究逐渐增多,如Hu等[9]利用来自水稻、小麦、燕麦和大麦的46个多态性微卫星标记对118份来自不同生态区的糜子资源进行了遗传多样性评价,将这些资源聚为5个类群;Hunt等[10]运用16个从糜子开发的微卫星标记分析了33份中国材料和来源于欧亚大陆的65份材料的遗传多样性。但有关糜子矮秆资源发掘的研究极少。张博等[11]研究表明,外施赤霉素可以使糜子矮秆突变体778恢复到原始高秆材料260(石湖千斤糜)的株高,推测导致突变的基因可能是赤霉素合成途径基因,这是有关糜子矮秆研究的唯一报道。
本研究利用EMS化学试剂诱导得到了1个新的糜子矮秆突变体“819”,通过将该突变体与高秆材料J12杂交产生F2代分离群体,系统调查亲本“819”、J12及其F2代群体株高、穗部相关性状,旨在确定矮秆基因的遗传特性以及10个性状间可能存在的连锁关系,为后续定位矮秆突变基因提供理论依据。
1 材料与方法
1.1 材料
试验所用的矮秆突变体“819”由糜子高秆材料“石湖千斤糜”经过EMS(甲基磺酸乙酯)诱变处理后获得,再用“819”与亲缘关系较远的高秆材料J12杂交获得674个F2代糜子植株个体。
1.2 试验设计
2017年在河北省张家口市农业科学院试验站(张家口市沙岭子镇)开始进行试验。观察土壤湿润度,在土壤湿润度适宜(相对湿度80%)的情况下旋耕土地;平整土地后6月17日进行材料播种,株距10 cm,行距33 cm;待6月23日糜子出苗后对其进行间苗、培土、除草,整个生育期未施肥;糜子抽穗前进行灌水1次,中耕除草2次。在试验田四周搭建防护网实行鸟害防治,未喷洒农药进行病虫害治理。
1.3 性状调查
糜子基本成熟后,对糜子杂交亲本及F2代群体的每一株单独进行田间性状的测定。测定株高、穗长、穗颈长、分蘖数、茎节数、二级枝梗长度、二级枝梗间距等10个性状。
株高:挑选每一株糜子中最高的主穗,用测量尺从糜子植株地表以上开始测量直至该糜子主穗穗尖的高度(cm)。通过绘制F2代群体株高分布密度直方图确定高秆、矮秆判定标准;如果呈现双峰分布模式,则以2个峰间分布频次最低的株高作为分界线,低于此株高判定为矮秆植株,高于此株高判定为高秆植株。
穗长:用短测量尺测定穗基部到穗顶的长度(cm)。
穗颈长:用短测量尺测定穗基部到最接近茎节的长度(cm)。
分蘖数:从糜子植株的根部对糜子的分蘖进行计数。
茎节数:从根部往上对糜子的茎节进行计数(节)。
二级枝梗长度1:从穗基部开始测定最近的一个糜子主穗的分支长度(cm)。
二级枝梗长度2:测定距离二级枝梗长度1最近的糜子分支长度(cm)。
二级枝梗长度3:测定距离二级枝梗长度2最近的糜子分支长度(cm)。
二级枝梗间距1:测量二级枝梗长度1与二级枝梗长度2之间的距离(cm)。
二级枝梗间距2:测量二级枝梗长度2与二级枝梗长度3之间的距离(cm)。
1.4 数据统计
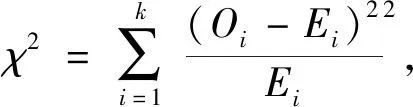
2 结果与分析
2.1 矮秆突变体819和J12双亲株型性状比较
矮秆突变体819和J12的株型性状比较见图1。父本J12的株高、穗长、穗颈长、茎节数、二级枝梗长度1、二级枝梗长度2、二级枝梗长度3极显著高于矮秆突变体819(P<0.01)。父本J12和矮秆突变体819的分蘖数、二级枝梗间距1和二级枝梗间距2无显著差异(P>0.05)。
2.2 糜子F2群体的农艺性状
糜子F2群体的株高平均值为108.455 cm,变异系数为22.6%,株高群体变异幅度很大,峰度与偏度都小于1,说明株高性状在F2群体上呈连续分布。糜子F2群体的穗长平均值为23.415 cm,穗长的变异系数是26.7%。糜子F2群体穗颈长介于2~34 cm,平均穗颈长为17.693 cm,变异系数是26.3%。分蘖数为0~16,极差值为16,分蘖数的平均值是6.630,变异系数是34.0%。茎节数变幅是4~13,全距为9节,平均茎节数7.310节,茎节数的变异系数为15.1%。二级枝梗长度1、二级枝梗长度2、二级枝梗长度3的变化范围分别是1.0~28.0 cm、4.0~29.0、1.7~29.0 cm,极差值分别为27.0、25.0、27.3 cm,变异系数分别是29.6%、28.4%、28.8%。糜子F2群体在二级枝梗间距1上的变化范围是0~7.2 cm,极差值是7.2 cm,变异系数是58.5%。
2.3 糜子F2群体的卡方检验
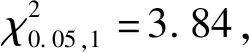
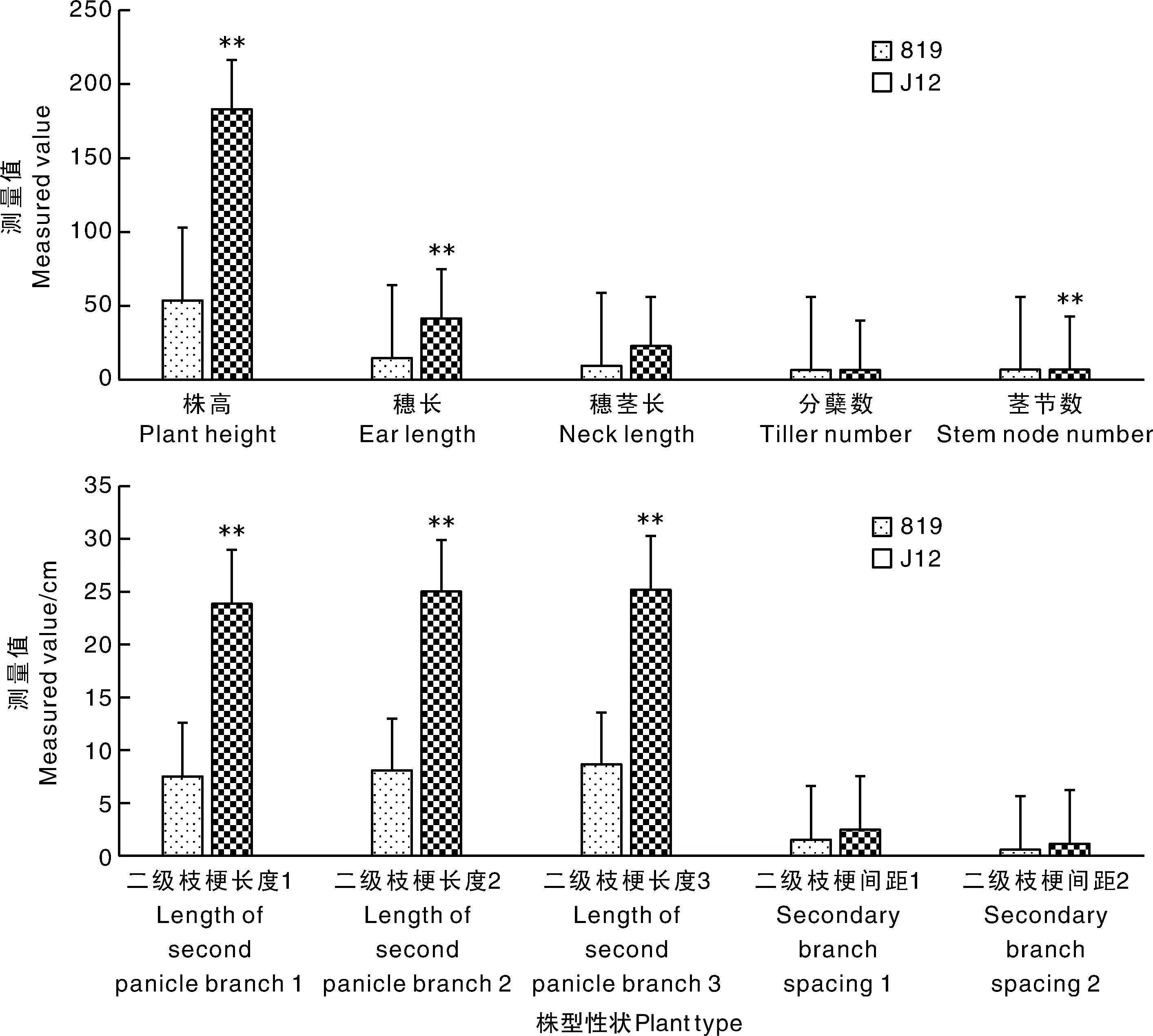
**表示差异极显著(P<0.01)。** meant significant difference at the levels of P<0.01.图1 矮秆突变体819和J12的株型性状比较Fig.1 Comparison of plant type traits between dwarf mutant 819 and J12
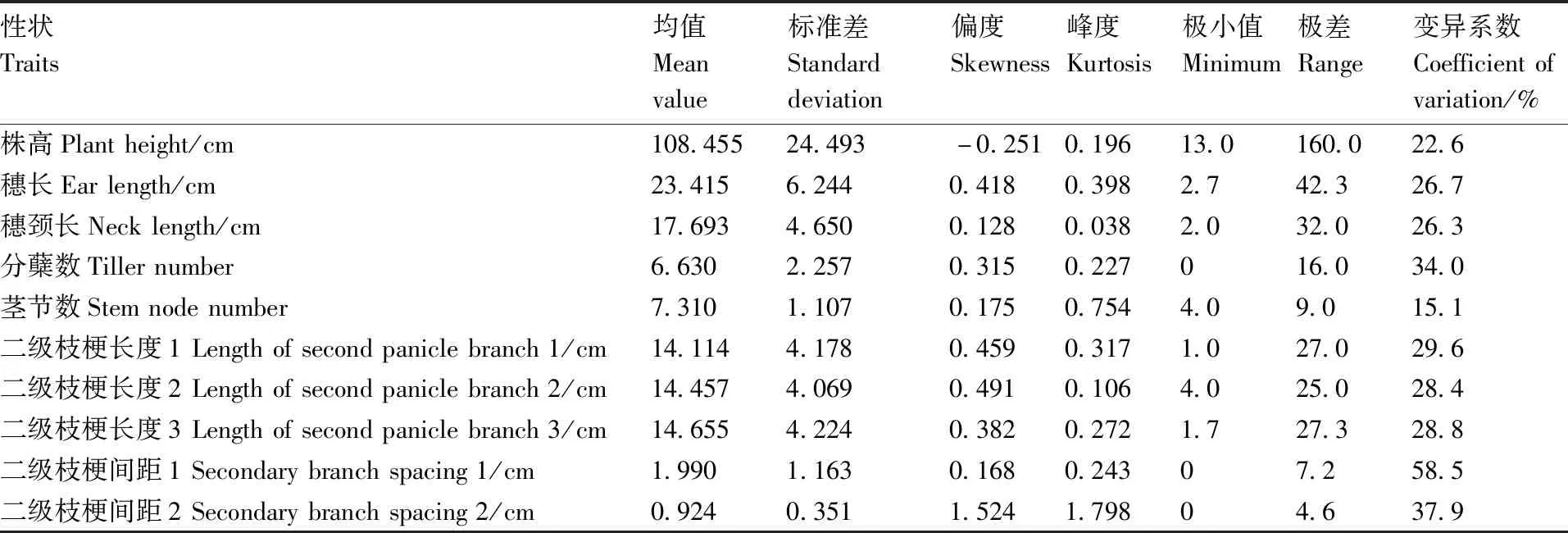
表1 F2群体的描述性统计分析Table 1 Descriptive statistical analysis of F2 population

图2 糜子F2群体的株型性状的频率分布图Fig.2 Frequency distribution of plant type characters in F2 population of millet
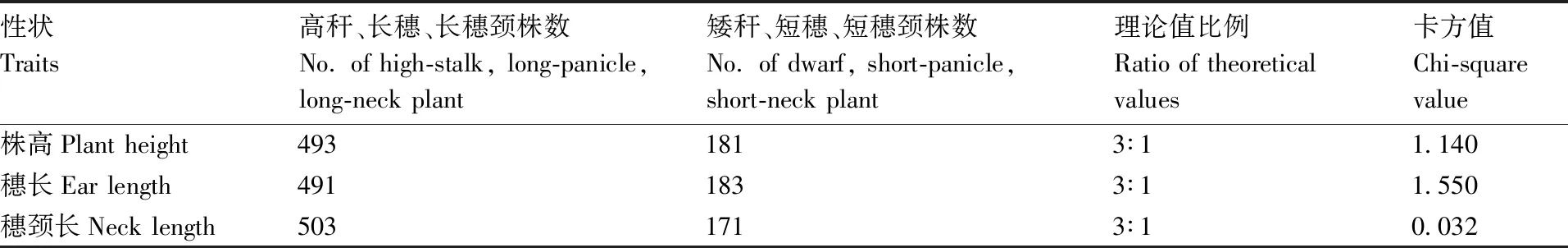
表2 卡方检验Table 2 Chi-square test
2.4 糜子性状间的相关与回归分析
糜子F2群体性状间相关分析见表3。糜子F2群体的株高与穗长、穗颈长、分蘖数、茎节数、二级枝梗长度1、二级枝梗长度2、二级枝梗长度3、二级枝梗间距1呈极显著正相关(P<0.01),株高与二级枝梗间距2呈显著正相关(P<0.05),其中株高与穗长(0.753)、二级枝梗长度1(0.705)、二级枝梗长度2(0.723)、二级枝梗长度3(0.725)的相关系数均高于0.5,为强正相关。穗长与穗颈长、茎节数、二级枝梗长度1、二级枝梗长度2、二级枝梗长度3、二级枝梗间距1、二级枝梗间距2呈极显著正相关(P<0.01),穗长与分蘖数呈显著正相关(P<0.05),其中,穗长与二级枝梗长度1(0.804)、二级枝梗长度2(0.831)、二级枝梗长度(0.831)的相关系数均高于0.5,为强正相关。穗颈长与二级枝梗长度1、二级枝梗长度2、二级枝梗长度3、二级枝梗间距1、二级枝梗间距2呈极显著正相关(P<0.01)。茎节数与二级枝梗长度1、二级枝梗长度2、二级枝梗长度3呈极显著正相关(P<0.01),与二级枝梗间距1呈极显著负相关(P<0.01)。二级枝梗长度1、二级枝梗长度2、二级枝梗长度3这3个性状两两间呈极显著正相关。糜子F2群体的株高(Y)对穗长(X1)、穗颈长(X2)、分蘖数(X3)、茎节数(X4)、二级枝梗长度1(X5)、二级枝梗长度2(X6)、二级枝梗长度3(X7)、二级枝梗间距1(X8)、二级枝梗间距2(X9)进行逐步回归分析,获得株高(Y)对其他农艺性状的回归方程为Y=-18.446+1.491X1+1.222X2+6.827X4+1.319X7+0.746X8,回归方程的拟合度为0.811,回归方程经显著性检验达到极显著水平(P<0.01)。表明可以利用此回归方程对糜子株高进行预测。

表3 F2群体的株型性状相关性分析Table 3 Correlation analysis of plant type traits in F2 population
**表示在0.01水平(双侧)显著相关,*表示在0.05水平(双侧)显著相关。
** indicated significant correlation at 0.01 level (bilateral); * indicated significant correlation at 0.05 level (bilateral).
2.5 糜子株型性状的主成分分析
糜子F2群体10个性状的主成分分析和主成分的因子得分结果见表4。由表4可以看出,特征值大于1的有3个主成分,这3个主成分的累积贡献率高达72.656%,即这3个主成分所涵盖的信息量可以反映出原来10个性状72.656%的信息特征。在第1主成分上,株高、穗长、二级枝梗长度1、二级枝梗长度2、二级枝梗长度3的因子得分比较高,分别为0.171、0.185、0.185、0.187、0.182,结合前面的相关分析可知这几个性状具有极显著正相关,因此第1主成分可以命名为长度因子。在第2主成分上,分蘖数、茎节数、穗颈长、二级枝梗间距1的因子得分比较高,分别为0.381、0.574、-0.281、-0.447,考虑到茎节数的因子得分比较高,因此第2主成分可以命名为茎节数因子。在第3主成分上,二级枝梗间距2的因子得分最高,为0.899,因此第3主成分可以命名为枝梗间距因子(表5)。
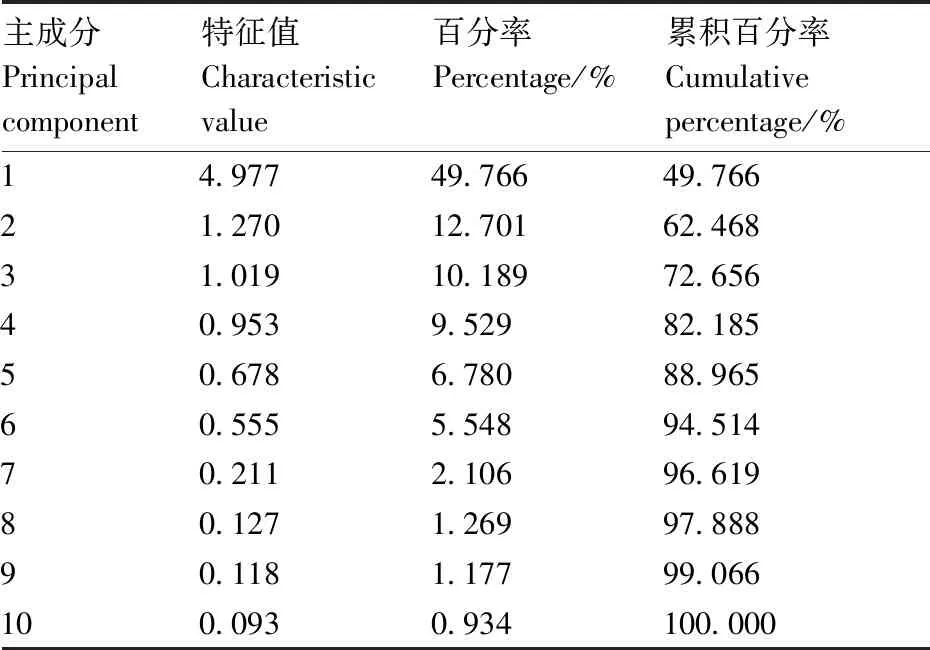
表4 糜子F2群体的株型性状的主成分分析Table 4 Principal component analysis of plant type characters in millet F2 population

表5 糜子F2群体株型性状主成分的因子得分Table 5 Factor score of plant type characters in principal component analysis of millet F2 population
3 讨论
作物的高矮直接影响到植株的生长状态、光合作用、种植密度、养分的吸收利用,最终影响到产量[12]。获得矮秆资源、培育矮化作物品种是增强抗倒伏性、增加种植密度从而提高产量的重要途径。通过一定条件筛选和特殊手段处理后可以得到比作物野生型对照株高明显降低的矮秆个体[13]。这些处理方法包括物理诱变、化学诱变和生物诱变3种,其中化学诱变广泛用于作物矮秆材料的创制[14]。目前,水稻、小麦、大麦、玉米等农作物已经通过化学诱变获得了大量矮秆突变体,已经运用到育种当中,对提高粮食产量做出了重要贡献。糜子是我国西北地区重要的杂粮作物,虽然利用传统杂交育种技术获得了一些优良品种[15],但是倒伏一直是影响糜子产量和品质的重要因素。发掘糜子矮秆资源是选育抗倒伏、高产糜子新品种的前提。
本研究利用EMS化学诱变方法获得了1个糜子矮秆突变体“819”,遗传分析表明突变体矮秆基因受隐性单基因控制。同时,在小麦、水稻、大豆、玉米矮秆突变体研究中均发现矮秆性状由隐性单基因控制[16-19],推测隐性遗传是矮秆性状较为普遍的一种遗传方式。事实上多数纯合矮秆材料由于植株过于矮小、穗小等原因都不能直接应用于育种实践,因此,本研究获得的矮秆突变体“819”还需要与合适的高秆材料进行杂交,改善穗部性状,适度增加株高,才能用于生产实践。本研究还发现,糜子分蘖数与株高呈极显著相关,而分蘖数会对糜子空间分布和种植密度产生直接影响,分蘖数的多少直接决定了是密植型还是稀植型;糜子穗长与产量有着密切关系,长穗一般籽粒较稀疏,产量潜能反而不高,同时长穗会导致植株易倒伏。黄英杰等[20]的研究结果表明,株高对产量影响达到极显著水平,而穗长对产量的总效应和直接效应均为负值,因此,品种选育时穗过长不可取。随着糜子株高的增长,糜子穗颈长、茎节数、二级枝梗长度、二级枝梗间距增加,穗长和分蘖数也随之增加,理论上糜子产量也会增加。但是实际上由于糜子株高过高时会导致糜子植株的倒伏,且植株的增长对土壤肥力也是一种消耗,所以适当降低糜子的株高,使其抗倒伏性和分蘖数、穗籽粒密度达到理想的平衡,可能会实现高产。
