47份外引小麦种质中矮秆基因的检测及其降秆效应分析
2021-09-23李怡鑫陈向东张雪宁林孟杰轩庆庆姜小苓王润发茹振钢吴晓军
李怡鑫,陈向东,张雪宁,林孟杰,尤 丽,李 冉,轩庆庆,姜小苓,姜 豪,王润发,茹振钢,吴晓军
(河南科技学院小麦中心/河南省杂交小麦重点实验室/现代生物育种河南省协同创新中心,河南新乡 453003)
提高小麦产量对于全球粮食安全至关重要。株高是小麦重要农艺性状之一,对粮食产量和品质影响较大。20世纪60年代的“绿色革命”,育种工作者将矮秆或半矮秆基因引入小麦,显著提高了抗倒伏能力和粮食收获指数,小麦产量也得到大幅提高[1-2]。
迄今为止,被鉴定的小麦Rht矮秆基因共有25个[3-4]。根据对外源赤霉素的反应,这些基因又被分为赤霉素敏感型和赤霉素不敏感型。对赤霉素不敏感的矮秆基因Rht-B1b(Rht1)和Rht-D1b(Rht2),降低株高的能力相当,一般能达到24%左右[5-6];对赤霉素敏感的矮秆基因,如Rht4、Rht5、Rht8、Rht9、Rht12和Rht13,通过缩短不同节间的长度,使株高分别降低45%、50%、11%、24%、46%和24%[7-11]。Rht-B1b和Rht-D1b来源于日本农林10号,Rht8来源于日本赤小麦[12-14]。一段时间以来,在我国小麦育种中应用较为广泛的矮秆基因主要有Rht-B1b、Rht-D1b和Rht8[15-17]。这种情况造成了国内矮秆基因利用单一化和小麦品种遗传基础日益狭窄,不利于产量潜力的进一步提高和小麦生产的可持续发展。另外,在小麦育种中已经被广泛利用的Rht矮秆基因,虽然具有较强的抗倒伏能性和良好的肥水反应,也显著增加了粮食产量,但仍然存在一定缺点,如综合农艺性状差、生产上利用困难等。因此,需要排除对农艺性状起负效作用的不利矮秆基因,寻找含有特定优良矮秆基因且综合性状好的小麦新种质,不断拓宽小麦种质遗传资源。
为了解决国内矮秆基因利用单一化和拓宽小麦种质资源,本研究以47份外引小麦种质为材料,利用与8个矮秆基因(Rht-B1b、Rht-D1b、Rht4、Rht5、Rht8、Rht9、Rht12和Rht13)紧密连锁的分子标记进行检测,鉴定外引种质中矮秆基因的类型和分布特点,筛选出国内利用较少且含有优良矮秆基因的外引种质材料,以期为有效利用外引小麦种质和提高小麦育种成效提供一些理论依据和材料支撑。
1 材料与方法
1.1 供试材料
47份小麦种质分别引自智利、美国、墨西哥、澳大利亚等10多个国家,其中11份材料(Kanto107、Manital、Opata、Glenlea、Madsen、Attila、Salmone、RL6077、Pavon、Pastor、Norin61)由山东省农业科学院作物研究所资源库提供,其余材料由河南科技学院小麦中心提供。小麦品种中国春(China Spring,CS)不含本研究所检测的矮秆基因[18],作为对照。
1.2 方 法
1.2.1 田间小麦株高的测量
小麦材料分别种植于河南科技学院朗公庙试验田和辉县试验田,每个品种(系)种植2行,行长4 m,行距20 cm,株距10 cm,试验地肥力均一,采用常规田间管理。在小麦灌浆期至成熟期测量株高,每个材料随机测量5株,记录数据。
1.2.2 DNA提取及分子标记检测
采用CTAB法提取小麦基因组总DNA,将DNA稀释至100 mg·μL-1,置于4 ℃冰箱保存备用。所用检测引物参照Ellis等[19-20]和Korzun等[21],提供的引物序列由上海赛默飞世尔科技(中国)有限公司合成,引物信息详见表1。
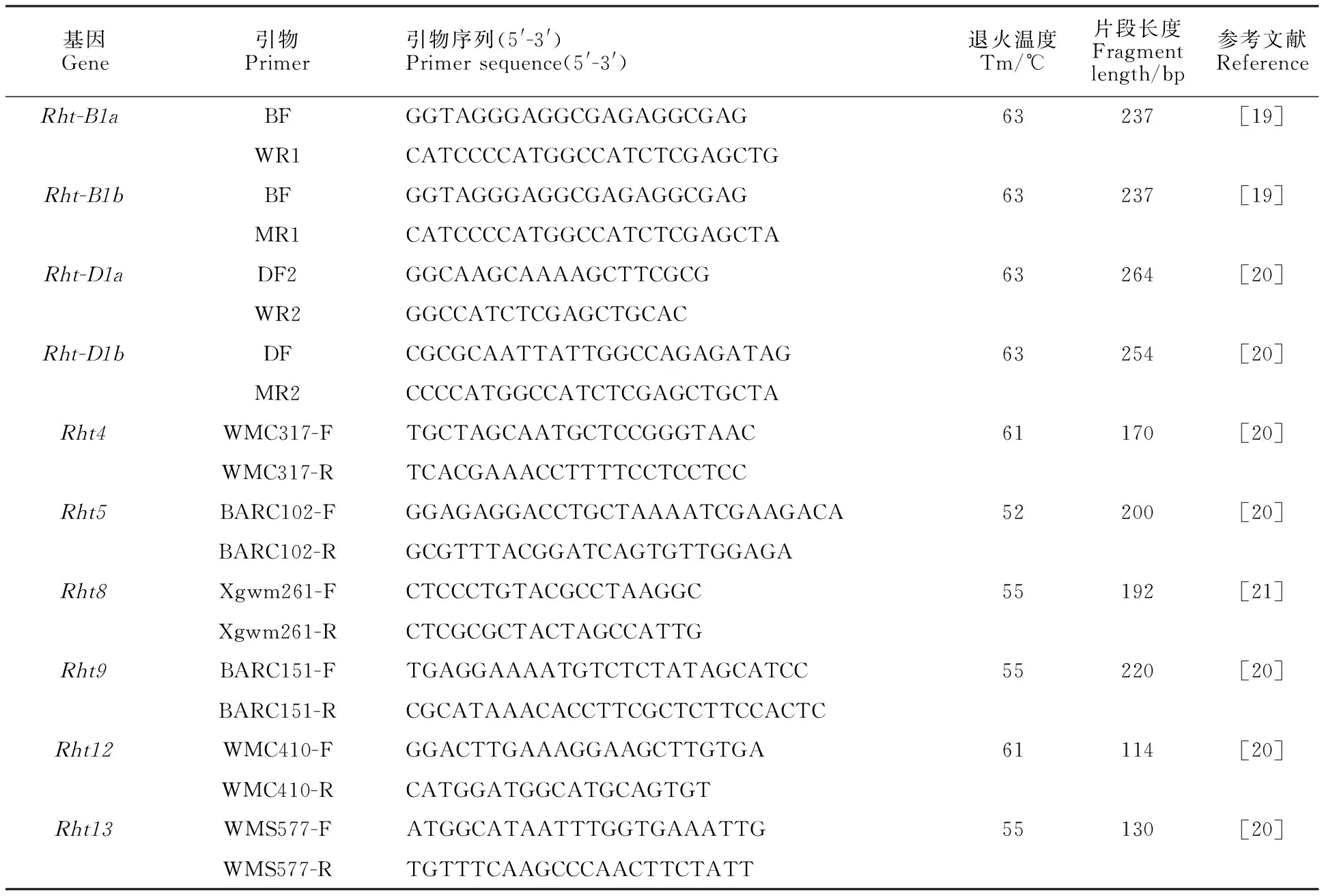
表1 矮秆基因引物信息Table 1 Primer information of dwarf genes
PCR扩增反应体系为20 μL,包含10 μL 2×Taq Master Mix(购至北京康为世纪生物科技有限公司)、10 μmol·L-1的正反向引物 0.6 μL、100 ng·μL-1的 DNA模板1 μL、ddH2O 7.8 μL。PCR扩增程序:95 ℃预变性5 min; 94 ℃变性40 s,52~63 ℃退火30 s,72 ℃延伸 1 min,35个循环;最后72 ℃延伸5 min。Rht1和Rht2扩增产物使用1.2%琼脂糖凝胶在1×TAE缓冲液中进行电泳检测;Rht4、Rht5、Rht9、Rht12和Rht13扩增产物使用2.0%琼脂糖凝胶进行电泳检测;Rht8扩增产物使用2.5%琼脂糖凝胶进行电泳检测,用凝胶成像系统拍照、保存。
1.3 数据统计分析
采用DPS 7.05进行差异显著性分析。
2 结果与分析
2.1 47份外引小麦种质中8个矮秆基因的分子标记检测
Rht1基因的多态性有两种,包括野生型(Rht-B1a)和突变型(Rht-B1b),其中,Rht-B1b表现为矮秆表型。利用两对引物BF/WR1和BF/MR1对供试材料进行检测,如果小麦种质中含有Rht-B1a基因,BF/WR1可以扩增出237 bp的条带,而BF/MR1无扩增条带;如果小麦种质种含有Rht-B1b基因,则BF/WR1无扩增条带,而BF/MR1可以扩增出237 bp的条带。本研究中,用不含所检测矮秆基因的小麦品种中国春为对照,BF/MR1引物扩增结果显示没有237 bp条带,说明扩增结果可靠(图1)。检测结果(表2)显示,47份外引材料中只含Rht-B1b单个矮秆基因的材料有2份,含有Rht-B1b或同时含有其他矮秆基因的材料有19份,占40.43%。
Rht2基因的野生型为Rht-D1a(264 bp),突变型为Rht-D1b(254 bp),Rht-D1b表现为矮秆表型。利用两对引物DF2/WR2和DF/MR2对供试材料进行检测,在含有Rht-D1a的小麦种质中,引物DF/MR2无扩增目的条带,而DF2/WR2则能扩增出264 bp的条带;在含有Rht-D1b的小麦种质中,DF/MR2可以扩增出254 bp的条带,而DF2/WR2无目的条带。47份外引小麦种质检测结果(图1,表2)显示,没有发现只含Rht-D1b单个矮秆基因的材料,同时含有Rht-D1b矮秆基因的材料有9份,占 19.15%。
根据Korzun等[21]设计的Rht8基因分子标记,利用引物Xgwm261-F/R对供试材料进行检测,如扩增出192 bp的目的条带,则表明小麦种质中含有Rht8矮秆基因。检测结果(图1,表2)显示,47份外引材料中,只含Rht8单个矮秆基因的材料有3份,含有Rht8或同时含有其他矮秆基因的材料有15份,占31.91%。
根据Ellis等[20]设计的Rht4、Rht5、Rht9、Rht12和Rht13基因分子标记,分别利用引物WMC317-F/R、BARC102-F/R、BARC151-F/R、WMC410-F/R和WMS577-F/R对供试材料进行检测。结果(图1,表2)显示,47份外引材料中,只含Rht4单个矮秆基因的材料有1份,含有Rht4或同时含有其他矮秆基因的材料有19份,占40.43%(图1、表2);只含Rht5单个矮秆基因的材料有2份,含有Rht5或同时含有其他矮秆基因的材料有4份,占8.51%(图1、表2);没有发现只含Rht9单个矮秆基因的材料,同时含有Rht9矮秆基因的材料有3份,占6.38%(图1、表2);只含Rht12单个矮秆基因的材料有2份,含有Rht12或同时含有其他矮秆基因的材料有19份,占40.43%(图1、表2);没有发现只含Rht13单个矮秆基因的材料,同时含有Rht13矮秆基因的材料有9份,占19.15%(图1、表2)。

M1:DNA marker DL2000;M2:50 bp DNA ladder;1:中国春;2:萨尔斯堡;3:澳阿优1号;4:阿大学AGT-②;5:KPL-1;6:鸟麦;7:KPL-6;8:KPL-7;9:KPL-11;10:Rudu。箭头所示为目标片段。
2.2 47份外引小麦种质中矮秆基因的分布及降秆效应
由表2和表3可知,47份外引小麦种质中有46份含有矮秆基因,占比接近98%。不同矮秆基因及其组合对小麦的降秆效应不同,单个矮秆基因的降秆效应不一定低于含有该基因的矮秆基因组合,不含矮秆基因的小麦材料株高显著高于含有矮秆基因的小麦材料。以不含8个矮秆基因的外引种质“非黑土地24”平均株高为标准,计算各矮秆基因及其组合的降秆效应,结果显示,降秆效应达到30%以上的矮秆基因组合有Rht-B1b/Rht8/Rht13(42.22%)、Rht4/Rht5(41.33%)、Rht-D1b/Rht4/Rht5/Rht12(38.37%)、Rht-B1b/Rht8(37.56%)、Rht-B1b/Rht4/Rht13(37.48%)、Rht-D1b/Rht4/Rht12(36.37%)、Rht-B1b/Rht4(35.41%)、Rht4/Rht12(34.22%)、Rht-B1b/Rht12/Rht13(33.26%)、Rht-B1b/Rht-D1b/Rht4/Rht12(32.44%)、Rht-B1b/Rht12(30.67%)、Rht4/Rht13(30.07%),8个矮秆基因中Rht5单个矮秆基因的降秆效应也达到了30%以上。
47份外引小麦种质中含有Rht-B1b、Rht-D1b、Rht4、Rht5、Rht8、Rht9、Rht12、Rht13单个矮秆基因的材料分别有2、0、1、2、3、0、2、0,共占总数的21.28%。含有2个及以上矮秆基因的材料占76.60%,其中含有2个矮秆基因的材料有23份,占48.94%,含有3个矮秆基因的材料有11份,占23.40%,含有4个矮秆基因的材料有2份,占4.26%。
47份外引小麦种质中8个矮秆基因的分布频率高低依次为Rht4=Rht12=Rht-B1b(40.43%)>Rht8(31.91%)>Rht-D1b(19.15%)=Rht13(19.15%)>Rht5(8.51%)>Rht9(6.38%)。含有这8个矮秆基因的小麦种质在株高上差异显著,降秆效应最低的为Rht9,平均株高101.07 cm;其次为Rht8,平均株高 95.67 cm,Rht12和Rht13降秆效应差异较小,分别为92.87 cm和92.75 cm,Rht-B1b、Rht-D1b、Rht4的降秆效应差异较小,分别为90.69 cm、 91.45 cm、91.98 cm,降秆效应最高的为Rht5,平均株高为86.65 cm(表2,表3)。

表2 8种矮秆基因及其组合的对株高的影响Table 2 Effect of eight dwarf genes and their combinations on plant height

表3 47份外引种质中矮秆基因类型及株高表现Table 3 Dwarf genotypes and plant height performance of 47 introduced germplasms
3 讨 论
矮秆或半矮秆基因应用于小麦育种,能够提高小麦的抗倒性和收获指数,对于提高小麦产量具有重要意义[22-24]。一直以来,我国小麦育种围绕少数骨干亲本来培育新品种,造成小麦遗传基础狭窄,也加大了培育突破性品种的难度。程西永等[25]对来自中国、智利、俄罗斯、墨西哥、澳大利亚、荷兰的728份小麦种质进行遗传多样性分析,发现中国小麦种质资源具有早熟、高粒重和植株矮的优点,而俄罗斯小麦分蘖成穗多,智利小麦穗粒数高,墨西哥小麦面粉吸水率高,荷兰小麦抗白粉病性好,说明从外国引进的这些小麦种质资源可以有效改良我国小麦的农艺性状、品质性状和抗病性。本研究中47份小麦种质分别引自墨西哥、智利、澳大利亚、美国、日本、俄罗斯等多个国家,抗病性遗传类型丰富,含有抗赤霉病、抗锈病、抗白粉病等基因,如澳阿优1号和bermude兼具条锈病和白粉病抗性[26]。其中,含有优异矮秆,或其他优良农艺性状基因的种质材料,可以适当应用到我国小麦育种中。
钟明志等[27]研究表明,Rht-B1b、Rht-D1b、Rht9和Rht13的降秆能力相当,降秆效应均在24%左右,其他矮秆基因的降秆能力表现为Rht5(50%)>Rht12(46%)>Rht4(45%)>Rht8(11%)。本研究中单个矮秆基因的降秆效应以Rht5最强,为31.78%,也是单个矮秆基因里唯一降秆效应达到30%以上的。李杏普等[22]研究表明,Rht12基因在显著降低株高的同时会促进小穗发育和提高穗粒数,也会造成生物产量降低、成熟晚等,但在以降秆为主要目的的育种中利用价值较大。本研究中含有Rht12单个矮秆基因的材料有2份,分别是Brennan和KPL-5,平均降秆效应只达到 18.96%,低于Rht8的降秆效应 (24.45%);有19份材料含有Rht12基因,占材料总数的40.43%,平均降秆效应为31.21%,远低于钟明志等[27]研究中Rht12的降秆效果。另外,还发现一些矮秆基因组合的降秆效应反而比单个矮秆基因弱,如Rht8单个矮秆基因的降秆效应为 24.45%,而Rht8/Rht12、Rht8/Rht13基因组合的降秆效应为18.67%和21.63%。造成以上现象的原因可能有三个,一是所检测小麦种质数量较少,二是除这8个矮秆基因外,还可能存在其他矮秆基因,三是株高为复杂的数量性状,受多种因素影响,引进的国外种质在新的生长环境中株高等性状表现 异常。
在矮秆基因应用方面,我国推广小麦品种含有的矮秆基因主要为Rht-B1b、Rht-D1b和Rht8等少数类型,而其他矮秆基因则利用较少[15-17]。除此之外,还有一些颇具应用潜力的矮秆基因,如Rht9在降秆的同时会减少小穗长度,增加分蘖数和千粒重,最终提高粮食产量[8];Rht12在显著降低株高的同时提高穗粒数,从而提高产量[10,22];Rht13对赤霉素敏感,在降低株高的同时不影响胚芽鞘长度和幼苗长势,而且可以显著提高穗粒数、增加粮食产量[24]。在本研究检测的47份外引种质材料中,未发现含有Rht9单个矮秆基因的材料,同时含有Rht9的材料共有3份,占材料总数的6.38%;未发现含有Rht13单个矮秆的材料,同时含有Rht13的材料共有9份,占材料总数的19.15%;含有Rht9、Rht12、Rht13的材料共有28份,占总数的59.57%,可以利用这些种质材料与国内品种组配,培育出广适性的小麦中间材料。47份外引种质出也含有降秆效果好而产量性状表现一般的矮秆基因,如Rht4、Rht5[27]。47份种质材料中,含有Rht4单个矮秆基因的材料有1份,含有Rht4或同时含有其他矮秆基因的材料有19份,占材料总数的40.43%;含有Rht5单个矮秆基因的材料有2份,含有Rht5或同时含有其他矮秆基因的材料有4份,占8.51%;其中,Rht4、Rht5与有应用潜力矮秆基因共存的材料有13份。47份外引种质矮秆基因类型和分布的明确,有利于在育种中排除某些农艺性状表现不良的矮秆基因(Rht4、Rht5),增加优良矮秆基因Rht9、Rht12、Rht13的育种利用,从而拓宽小麦矮秆育种遗传基础。另外,在8个矮秆基因中,除Rht12为显性遗传外,其余基因均为隐性遗传[27],而隐性遗传矮秆基因在小麦育种的早代进行选择相对困难,因此,利用分子标记进行早代辅助选择,有利于提高小麦育种效率。
本研究利用8个矮秆基因分子标记对47份外引种质材料的矮秆基因类型及分布频率进行了鉴定,明确了Rht-B1b、Rht-D1b和Rht8三个主要矮秆基因的分布频率,筛选出含Rht9、Rht12、Rht13有应用潜力矮秆基因的种质材料28份,为国内矮秆高产小麦品种选育提供了一些理论依据和材料支撑。
