天然珙桐种群结构与动态特征在高低纬度地区的差异
2020-03-05徐云飞刘沁松徐文娟唐俊峰董廷发杨宝臻
徐云飞 刘沁松 徐文娟 唐俊峰 董廷发 杨宝臻 胥 晓,3*
(1. 西南野生动植物资源保护教育部重点实验室,南充 637009;2. 西华师范大学生命科学学院,南充 637009;3. 西华师范大学西南山地特色植物种质适应与利用研究所,南充 637009)
珙桐(Davidia involucrata Baill.)是我国特有珍稀濒危植物,生存范围狭窄,仅分布在中国西南地区的四川、云南、贵州及周边省份,地理范围为27°~33°N、98°~111°E[1]。前人对珙桐的研究主要集中在珙桐的生物学、群落学、生理生化等方面[2~5]。近年来针对珙桐种群结构及其数量动态特征的研究引起了更多的关注[6~7]。如贵州柏箐喀斯特地区和四川都江堰龙池地区的珙桐种群结构均呈倒金字塔型,幼苗数量不足,为衰退型种群[8~9]。杨心兵等[10]对湖北省后河自然保护区光叶珙桐种群结构研究后发现光叶珙桐种群径级分布呈倒金字塔型,种群结构属于中衰型,而湖北星斗山自然保护区和甘肃文县的珙桐群落属于增长型种群[11~12]。
虽然上述研究调查了某个地区珙桐种群结构和数量动态特征,但珙桐种群特征在不同纬度区域上的差异尚未见关注。由于地表接收的太阳辐射随纬度的变化差异非常显著,导致地球上的热量分布随纬度升高而降低[13],同时,土壤积累的储水量与净降雨量随纬度增加也呈现降低的趋势[14]。因此,不考虑非地带性影响,不同纬度地区间温度、光照、降水等环境因素差异明显,这种差异将影响植物种群的径级结构和数量。如刘忠成[15]等研究发现大果马蹄荷(Exbucklandia tonkinensis)群落的重要值及种群径级结构大小与地理分布规律表现出一致性。甜锥(Castanopsis eyrei)在粤北6 地的个体数量分布与各地的纬度差异明显相关,处于较高纬度地区的个体数量较多,纬度较低的地区个体数量少[16]。红树林植物高度随纬度的增加变矮,生物量也随纬度增加而减少[17]。不同纬度的马尾松径向生长对气候变化的响应不同[18]。鉴于纬度对植物种群的影响主要表现在热量和降水,二者是影响植物分布的重要因子,而濒危植物与其生境的依赖关系远远高于其他植物,因此,我们推测不同纬度地区天然珙桐种群的种群结构和动态特征将具有显著差异。在中国境内,除塔里木和藏北高原,同一经度上的年均温和年降水量随纬度升高而递减[19],低纬度地区天然珙桐种群的热量和降水将高于高纬度地区,因而低纬度比高纬度地区珙桐种群可以获得更充分的水热条件。为验证我们的推测,我们以四川平武县和云南滇东北地区的天然珙桐种群为研究对象,二者几乎在同一经度上,能避免由经度差异带来的影响,通过对其年龄结构、种群动态特征和生存力进行研究,预测珙桐种群对水热变化的响应规律,以此揭示纬度对天然珙桐种群结构和动态特征的影响。
1 研究区域
平武县珙桐种群样地位于四川省绵阳市境内。样地的地理坐标为32°19′N,104°32′E。海拔1 701~1 752 m,属亚热带山地湿润季风气候,年均温9.5℃,年均降水量957 mm(https://www.meteoblue.com),土壤属于黄壤类型。样地中乔木层以灯台树(Bothrocaryum controversum)、槲栎(Quercus aliena)为主,灌木层以马桑(Coriaria nepalensis)、火棘(Pyracantha fortuneana)为主。
滇东北珙桐种群位于云南省昭通市内。分布区 中 心 地 理 坐 标 为27°94'N,104°22'E,海 拔1 530~2 050 m,属中亚热带气候,年均温17.9℃,年降水量1 470 mm(https://www.meteoblue.com)。土壤属于黄壤类型。样地中乔木层以珙桐、峨眉栲(Castanopsis platyacantha)、水青树(Tetracentron sinensis)为主,灌木层以筇竹(Qiongzhuea tumidissinoda)、绥江玉山竹(Yushania suijiangensis)、金佛山方竹(Chimonobambusa utilis)为主[20]。
2 研究方法
2.1 数据来源
在对平武县药丛山珙桐种群全面踏查的基础上,选择典型地段设置3 个样地,样地面积约700 m2,对样地内珙桐个体进行每木调查,记录株高、胸径、冠幅等,用手持GPS定位仪记录经纬度、海拔等,同时记录调查时间、坡度、坡向、郁闭度等环境因子,各样地信息基本信息见表1。滇东北珙桐种群的样地情况和统计的径级结构数据引用自罗柏青等[20],珙桐种群地理位置见图1。基于高低纬度珙桐种群的径级结构数据编制静态生命表,绘制存活曲线、死亡率和消失率曲线以及生存函数曲线,预测未来种群结构特征。

表1 各样地基本信息Table 1 The basic information of the sample plots
2.2 龄级划分
珙桐的胸径生长与年龄呈显著的正相关,因此可用胸径大小来推测珙桐的树龄[21]。本研究采用径级为划分龄级结构的标准,以此分析珙桐种群的年龄结构特征[22]。根据珙桐生长特点,把平武县种群和滇东北种群的珙桐胸径(DBH,Diameter at Breast Height)的大小划分为11 个径级:Ⅰ,DBH<5 cm;Ⅱ,5 cm≤DBH<10cm;Ⅲ,10 cm≤DBH<15 cm;Ⅳ,15 cm≤DBH<20cm;Ⅴ,20 cm≤DBH<25 cm;Ⅵ,25 cm≤DBH<30cm;Ⅶ,30 cm≤DBH<35 cm;Ⅷ,35 cm≤DBH<40 cm;Ⅸ,40≤DBH<45 cm;Ⅹ,45 cm≤DBH<50 cm,Ⅺ,50 cm≤DBH<55 cm。第Ⅰ径级对应第Ⅰ龄级,第Ⅱ径级对应第Ⅱ龄级,以此类推。统计各级珙桐个体数,并编制珙桐种群静态生命表。

图1 珙桐种群的地理位置Fig.1 The location of D.involucrata populations
2.3 种群动态量化方法
珙桐种群结构动态分析方法参考陈晓德[23]。计算公式如下:

式中,Sn和Sn+1分别为第n 和第n+1 年龄级的个体数,-1≤Vn<1。Vn为种群从n到n+1级的个体数量变化。Vpi为忽略外部干扰时整个种群结构的数量动态变化指数,当考虑外部干扰时可将公式进一步修正为:

式中:K 为种群年龄级数量,Vn、Vpi、V'pi值为正、负、零时分别反映种群个体数量的增长、衰退、稳定的结构关系。
2.4 种群静态生命表和生存规律分析
静态生命表的编制方法参见金慧等[24]。由于静态生命表仅反映了种群年龄动态历程中的一段特定时间,不是对种群整个生活史的追踪[25],在编制生命表时会出现死亡率为负的情况[26],因此本文采取匀滑技术处理[27],Ax即为ax匀滑修正后得到的数据。
珙桐种群的动态变化方法参考杨凤翔等[28]引入生存分析中的4个函数,计算公式如下:

2.5 种群数量动态的时间序列分析
本研究采用时间序列分析中的一次移动平均法 对 珙 桐 种 群 的 年 龄 结 构 进 行 预 测[29~30]。公式为:
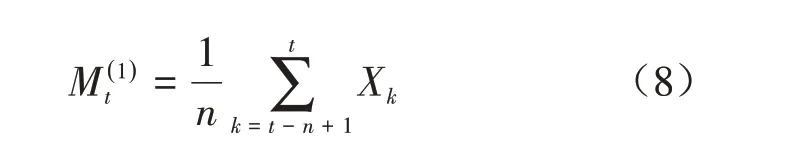
式中:n表示预测的时间(本研究为龄级),t为龄级,Xk为k龄级内的个体数量,M(1)t表示经过未来n个龄级时间后t龄级的种群大小。本研究对平武县未来经过2、4、6 个龄级和滇东北2、4、6、8 和10龄级时间后的种群各龄级的个体数量进行预测。

图2 珙桐种群年龄结构A. 平武县(高纬度);B. 滇东北(低纬度)(引自罗柏青等,2012) 下同。Fig.2 Age structure of D.involucrata populationsA.Pingwu(High latitude);B.Northeastern Yunnan(Low latitude)(Figure was derived from Luo,et al,2012) The same as below.
3 结果与分析
3.1 珙桐种群分布现状及结构特征
高纬度的平武县和低纬度滇东北珙桐种群结构均呈不规则的金字塔型(见图2),平武县种群主要集中在第Ⅱ龄级,占总数的36.17%,其次是第Ⅰ龄级,占总数的15.96%。平武县种群第Ⅶ、Ⅸ、Ⅹ和Ⅺ龄级缺失,其余龄级均有分布。处于低纬度的滇东北珙桐种群第Ⅱ龄级最多,占总数的32.5%,其次是第Ⅲ龄级,占总数的20.62%,第Ⅰ龄级占比为11.88%。滇东北种群各龄级均有一定数量的个体分布,种群结构稳定。
表2 显示,高纬度的平武县珙桐种群V1和V7都小于0,说明第Ⅰ和Ⅶ龄级均呈现衰退的结构动态关系,V2~V4和V6均大于0,表明第Ⅱ~Ⅳ和第Ⅵ龄级呈现增长的结构动态关系,V5等于0,说明在第V 龄级呈现稳定的结构动态关系;低纬度的滇东北珙桐种群V1、V7、V9均小于0,其余龄级均大于0,说明Ⅰ、Ⅶ、Ⅸ这3 个龄级更新较差,呈衰退的结构动态关系,其余龄级呈现增长的结构动态关系。
种群总体动态变化指数分析结果表明,高纬度地区的平武县和低纬度的滇东北珙桐种群Vpi值均大于0,分别为18.47%和25.51%,说明两地区珙桐种群同属于增长型种群,低纬度的滇东北种群比高纬度的平武县种群有更高的增长趋势。两地的V'i值分别为0和2.32%,在考虑外部干扰条件下高纬度的平武县种群受外部干扰能力低于低纬度的滇东北种群。
3.2 静态生命表和存活曲线
珙桐种群静态生命表如表3 所示,两地种群存活数(lx)随龄级增加逐渐减少。高纬度的平武县种群个体的期望寿命(ex)在Ⅰ、Ⅱ级后逐渐降低,低纬度的滇东北珙桐种群个体期望寿命(ex)随龄级先增加后减少,在第Ⅴ龄级达到最大值。

表2 珙桐种群动态变化指数Table 2 Dynamic indices of D.involucrata population

表3 珙桐种群静态生命表Table 3 Static life table of D.involucrata population

表4 珙桐种群存活曲线的检验模型Table 4 Test models of survival curves of D.involucrata population
以径级相对龄级为x 轴,以lnlx、qx、kx为y 轴绘制平武县和滇东北珙桐种群的存活曲线(见图3)、死亡曲线和消失率曲线(见图4)。存活曲线是反映种群个体在各年龄级的存活状况曲线,采用Hett 和Loucks[31]提出的两种数学模型(见表4)即指数方程式NX= N0e-bx和幂函数方程式NX=对存活曲线进行检验,结果显示,指数函数模型的R2值均大于幂函数模型,因此两地珙桐存活曲线趋近于Deevey-Ⅱ型。
由图4 可知,高纬度的平武县种群的死亡率和消失率曲线变化趋势基本一致,Ⅰ~Ⅳ龄级死亡率和消失率曲线增长趋势平缓,在第Ⅴ龄级后死亡率和消失率骤升。低纬度的滇东北种群Ⅰ~Ⅳ龄级死亡率和消失率曲线急剧增长,第Ⅴ~Ⅷ龄级增长趋缓,死亡率较高,中龄级个体受环境压力较小,生长状况更加稳定。
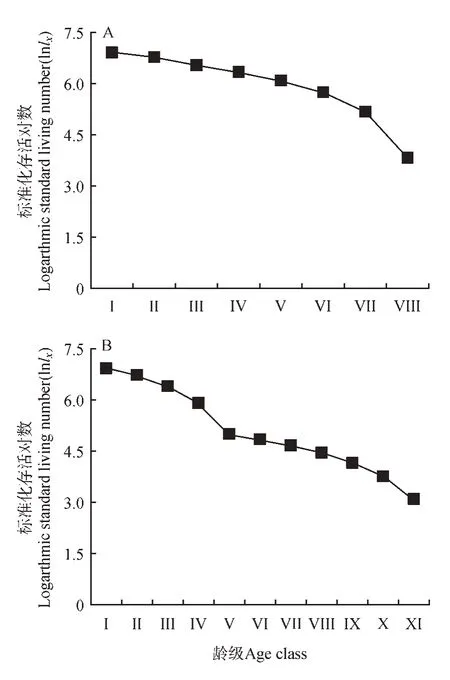
图3 珙桐种群的存活曲线Fig.3 Survival curves of D.involucrata population
3.3 珙桐种群的生存力
以龄级为横坐标,分别以生存函数的4 个函数值为纵坐标,分别绘制生存率曲线、累计死亡率曲线、死亡密度曲线和危险率曲线,如图5~6。
生存分析表明,两地区珙桐种群随龄级增加呈现出生存率和累计死亡率互补,死亡密度和危险率曲线基本一致的趋势。高纬度的平武县珙桐种群随龄级增加,生存率下降,累计死亡率上升,中龄级个体危险率逐渐升高;低纬度的滇东北珙桐种群在第Ⅰ~Ⅳ龄级生存率锐减,累计死亡率骤升,在第Ⅳ龄级后生存率和累计死亡率降低和增长的趋势较平缓。
平武县珙桐种群死亡密度在第Ⅰ~Ⅲ龄级略有波动,整体上呈现出“先增长再降低后增长”的趋势,危险率先增后减,在第Ⅴ龄级危险率最高;滇东北珙桐种群死亡密度和危险率在第Ⅰ~Ⅳ龄级波动较大,死亡密度在Ⅳ~Ⅷ龄级趋于稳定,危险率曲线随龄级逐渐上升,这与生存率和累计死亡率的结果一致,滇东北珙桐种群具有前期减少,中期稳定,后期衰退的特点。
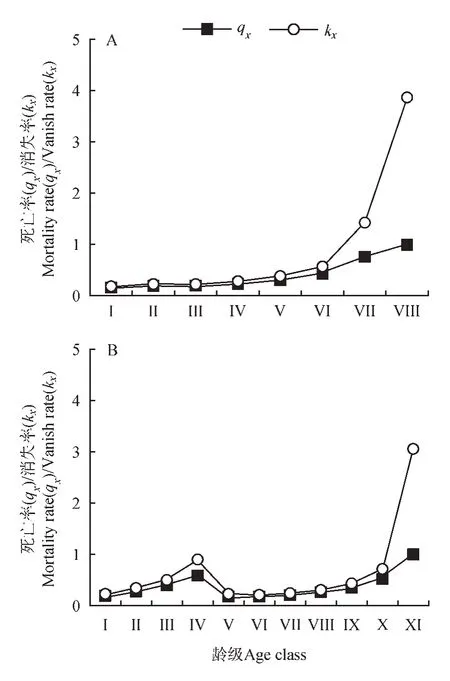
图4 珙桐种群死亡率(qx)和消失率曲线(kx)Fig.4 Mortality(qx)and vanish(kx)rate curves of D.involucrata population
3.4 珙桐种群数量的时间序列分析
高纬度的平武县和低纬度的滇东北珙桐种群数量动态时间序列预测以各地种群各龄级株数为原始数据,按照一次移动平均法预测出平武种群各龄级在未来2、4、6 个龄级时间后的个体数和滇东北种群各龄级在未来2、4、6、8、10 个龄级时间后的个体数(见表5)。结果显示,平武县珙桐种群个体总数将从目前的94株下降至6个龄级后的37株,珙桐剩余率30.8%。滇东北珙桐种群幼龄级个体随时间变化逐渐减少,中龄级个体逐渐增加,6 个龄级时间后珙桐剩余量为49.38%,表明滇东北珙桐种群个体丰富,可以维持未来较长时间种群正常更新。
4 讨论

图5 珙桐种群生存率(S(i))和累计死亡率曲线(F(i))Fig.5 Survival rate(S(i))and accumulated mortality rate(F(i))curves of D.involucrata population

图6 珙桐种群死亡密度(f(i))和危险率曲线(λ(i))Fig.6 Morality density(f(i))and hazard rate(λ(i))curves of D.involucrata population
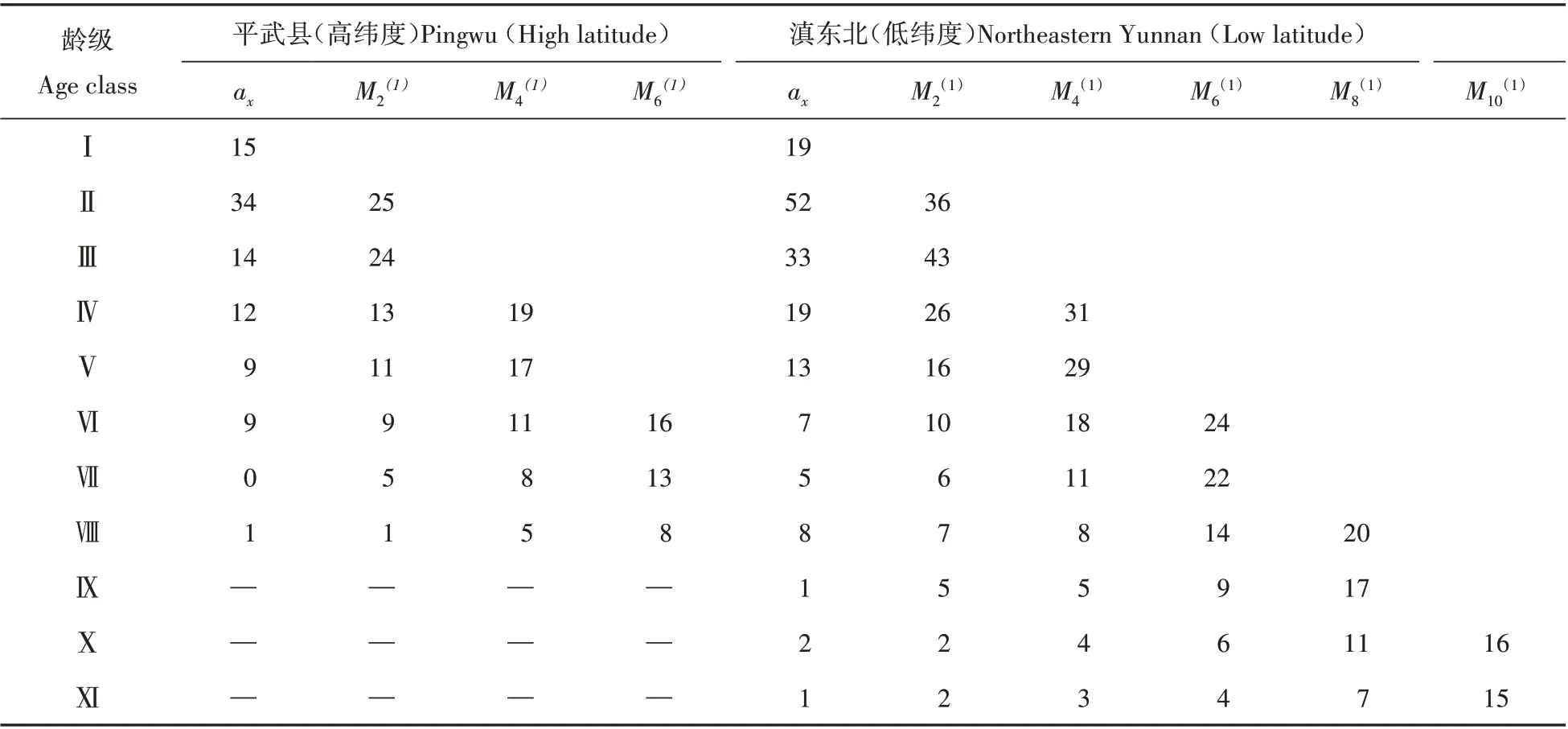
表5 珙桐种群动态变化的时间序列分析Table 5 Time sequence analysis of age structure of D.involucrata population
珙桐间断分布于我国长江流域的27°~33°N,98°~11°E,主要在四川省及周边的云南、湖北等地分布[32]。为了避免经度和土壤类型差异对本实验的影响,我们在珙桐分布区域的南北缘分别选取滇东北种群和平武县种群作为低、高纬度地区域的研究对象,纬度差为5°,经度差不足0.2°,二者的经度相差极小,土壤类型均为黄壤,两地珙桐种群同处在一个经度范围但分布在不同的纬度地区,温度和降水存在显著差异,两地珙桐种群结构与动态特征的差异可以客观地归结为纬度变化带来的影响。
研究表明,植物群落和种群受气候、土壤等因素强烈影响,环境差异不仅影响植物群落特征,同时对种群结构也有显著影响[33],比较不同生境下种群特征有助于揭示物种与其生存环境之间的适合度[34]。本研究发现高低纬度两地区的珙桐种群年龄结构均呈现不规则的金字塔型。从种群大小来看,滇东北种群在个体数量上优于平武县种群。从年龄结构来看,两地珙桐种群均呈现出幼龄级个体缺乏的特点,这与马宇飞[35]的研究结果一致。其中,Ⅰ、Ⅱ龄级个体在平武县种群中所占比例高于滇东北种群所占比例。结合动态指数来看,高纬度的平武县和低纬度的滇东北珙桐种群的第Ⅰ龄级动态指数分别为-55.88%和-63.46%,整体上滇东北珙桐种群幼苗衰退程度比平武县种群更高。因此,不同纬度引起水分条件的差异是导致幼龄级个体占比不同的主要因素,推测原因一方面是由于滇东北种群所处纬度低、降水丰富,但大量幼苗由于过湿或其他不利的环境胁迫导致幼苗死亡较多,最终经过发育进入乔木层的个体数量不多[36],另一方面滇东北珙桐生境中灌木层均以不同的散生竹类占绝对优势[20],珙桐幼树对地上和地下空间利用竞争能力弱,受环境筛选作用强烈[37],光照对珙桐幼苗和幼树生长影响显著[38]。结合当地生境条件,高纬度地区的平武县珙桐种群林下灌木、草本植物稀疏,为当地珙桐幼树的生长提供了光照和生长空间,这一结果与不同纬度的香果树(Emmenopterys henryi)种群结构的研究结果相似[39]。
种群静态生命表和存活曲线的分析结果表明,高纬度的平武县和低纬度的滇东北珙桐种群的存活曲线均趋向于Deevey-Ⅱ型。高纬度的平武县种群Ⅰ~Ⅳ龄级的死亡率曲线趋势相对稳定,但是Ⅳ龄级后死亡率急剧上升;低纬度的滇东北种群幼龄级个体死亡率略有波动,但中龄级个体死亡率和消失率逐渐上升但趋势平稳。表明平武县种群中龄级个体生长不利,原因可能是平武县种群珙桐幼龄级数量较多,在向成树发展过程中个体生长所需空间和营养物质增大,植株间竞争激烈,自疏作用更加明显,导致死亡率逐渐增高[40],珙桐受到的竞争强度随林木径级的增大而逐渐减少[41]。滇东北种群中龄级个体死亡率和消失率曲线趋势平缓原因可能是因高龄级个体的生理衰老所致。此外,生存分析结果表明,高纬度的平武县种群随龄级增加生存率下降,累计死亡率上升,中龄级个体危险率逐渐升高,低纬度的滇东北种群幼龄级个体累计死亡率高,中龄级个体危险率曲线趋势平缓,这与死亡率曲线结果一致。时间序列分析结果表明,两珙桐种群个体数随龄级增加种群数量下降,老龄个体逐渐增多,幼龄级个体更显不足。可以推测,如不加强对幼苗、幼树的保护,两地珙桐种群将难以保持稳定增长。尽管我们的上述结果已经可以证实高低纬度的珙桐种群在种群结构与动态特征方面存在显著差异,但两地的少数环境因子(如坡度和坡向等)对分析结果产生的误差还需要进一步分析和研究。
综上,天然珙桐种群的结构与动态特征存在纬度差异,高纬度的平武县珙桐种群群落幼龄级个体生长状况良好,但随龄级增加,自疏作用逐渐增强限制了中龄级个体生存,而低纬度的滇东北珙桐种群林下条件不适宜导致幼龄级更新不畅。从种群整体角度来看,低纬度种群在种群数量、生长趋势方面要优于高纬度种群。因此,对不同纬度的珙桐种群的保护应采取多种保护方法相结合的方式。高纬度的平武县珙桐种群可以采取人工补苗措施将实生幼苗栽回原种群以补充幼龄个体,并对中龄级个体进行适度的修剪甚至间伐以降低林间郁闭度。而低纬度的滇东北珙桐种群则要加强人工抚育,同时伐去母树和幼苗周围的灌木和草本植物,提高幼苗的成活率,增加幼苗密度。
