突脉金丝桃的花器官发生及其系统学意义
2020-03-05吴海燕
赵 祥 苏 雪 吴海燕 张 辉 孙 坤
(西北师范大学生命科学学院,兰州 730070)
金丝桃属(Hypericum)是被子植物中最大的100 个属之一[1],全世界约有496 种[2],除南北两极地或荒漠地及大部分热带低地外,在世界各地广布[3]。该属植物花最主要的特征为雄蕊多数,联合成束[3]。近年来有关金丝桃属分类地位的研究出现了不同的观点,传统的观点认为金丝桃属隶属于广义藤黄科(Clusiaceae(Guttiferae))金丝桃亚科(Hypericoideae)[3],而现有的分子系统学研究表明,广义的藤黄科并不是一个单系[4],因此APGⅢ系统将金丝桃属所属的金丝桃亚科从藤黄科中分出,独立为金丝桃科(Hypericaceae)[5]。根据分子系统学的研究,金丝桃科隶属于核心真双子叶植物蔷薇分支的金虎尾目[6]。
具有联合成束的雄蕊是金丝桃科的典型特征之一[7~9],然而这一特殊花部结构—雄蕊束的起源问题还存在着争议[10]。花部发育式样的研究以及不同类群某个性状演化系列的比较可以使我们更加准确地理解被子植物花的演化式样和推断类群间的演化关系[11],因此进行相关类群的花器官发生研究极为必要。目前,关于藤黄科[12~13]植物花器官发生的研究已有报道,但除早期的解剖学观察[14~15]外,金丝桃科植物的花器官发生发育仍缺乏研究。因此,为了揭示金丝桃科植物的花器官发育式样,探索其多数雄蕊及成束雄蕊结构的起源,本文以突脉金丝桃(Hypericum przewalskii)为研究材料,运用扫描电子显微镜对其花的形态发生和发育进行了观察,以期为从花器官发生的角度为揭示金丝桃科与藤黄科的系统关系提供证据。
1 材料和方法
研究材料采自甘肃省漳县,采集突脉金丝桃植株不同发育阶段的花芽及花蕾,置于FAA 固定液中固定保存,在室内经70%乙醇转入95%乙醇过夜后在解剖镜下将多余的花器官剥去,露出需观察的原基,经各级乙酸异戊酯梯度脱水处理,最后于真空CO2临界点干燥,喷金镀膜后置于HITACHIS-450扫描电镜下拍照观察[11]。
2 结果与分析
2.1 突脉金丝桃成熟花的结构
突脉金丝桃为两性花,花直径2~4 cm,开展,下方有2 枚叶状小苞片。萼片5。花瓣5,金黄色至柠檬黄色,长圆形。雄蕊多数,联合成不明显的5 束。合生雌蕊,子房卵球形,5 室,光滑;花柱5,自中部以上分离。
2.2 突脉金丝桃花部器官的发生
扫描电镜观察结果显示,2 枚小苞片最先发生,呈对生排列,大小几乎一样。花原基在苞片原基的包裹中完成分化过程,呈球形(图版Ⅰ:1~2)。萼片原基的发生是紧接着苞片原基的发育进行的,萼片原基以2/5 的圆周相继发生(图版Ⅰ:3~5)。位于与苞片互生位置的第1 枚萼片原基首先开始发生,紧接着第2 枚萼片在距第1 枚萼片2/5圆周的位置发生,随后第3 枚萼片在距第2 枚2/5圆周的位置发生,第4 枚、第5 枚以同样的距离和方向最后发生(图版Ⅰ:4~5)。萼片原基发生时呈隆起的椭球型,后渐渐发育成舌状,包裹花原基的顶部。
在萼片原基发生完成后,在花原基位于萼片原基之间角隅处形成花瓣-雄蕊共同原基(图版Ⅰ:5~6),5 枚雄蕊—花瓣共同原基近同时发生。共同原基发生后,其下部逐渐向外伸展,形成一个月牙形的花瓣原基,之后发育速度渐缓,上部向上形成凸起,形成与花瓣原基相对的雄蕊原基(图版Ⅰ:7)。花瓣发生时,5 枚花瓣原基没有明显的先后的顺序,几乎是同时发生的。5枚雄蕊原基的发生和花瓣原基的发生类似,几乎同时发生且无明显大小之分(图版Ⅰ:8~10)。随着雄蕊原基的向上延伸膨大,多数次生雄蕊原基开始由内向外依次发生(图版Ⅰ:11~12),发生方式为离心发生(图版Ⅱ:13~15)。随着次生雄蕊原基的发育和数目的增多,可以明显观察到5 束雄蕊的形成(图版Ⅱ:16~19,22)。
在次生雄蕊原基开始发生的同时,花原基顶端出现五个突起,心皮原基开始发生,原基之间的基部组织与突起一起发育使得每一心皮都在基部连成整体,而中部凹陷(图版Ⅰ:10)。之后由于心皮原基与基部组织的进一步向上延伸,心皮原基的凹陷逐渐加深使每一角呈V 字形状,心皮边缘沿着凹陷向中间靠拢,呈对折状,随着对折的加深,每一心皮原基的边缘逐渐愈合,发育为子房(图版Ⅰ:10~12,图版Ⅱ:13~19),而心皮顶端离生部分继续生长,形成花柱和柱头,在心皮还未完全愈合时就可观察到柱头乳突细胞的形成(图版Ⅱ:22~24)。在心皮原基进一步向上突起的过程中,次生雄蕊原基顶端出现人字形凹痕,逐渐分化出花药,而基部进一步延伸,形成花丝(图版Ⅱ:19~21)。
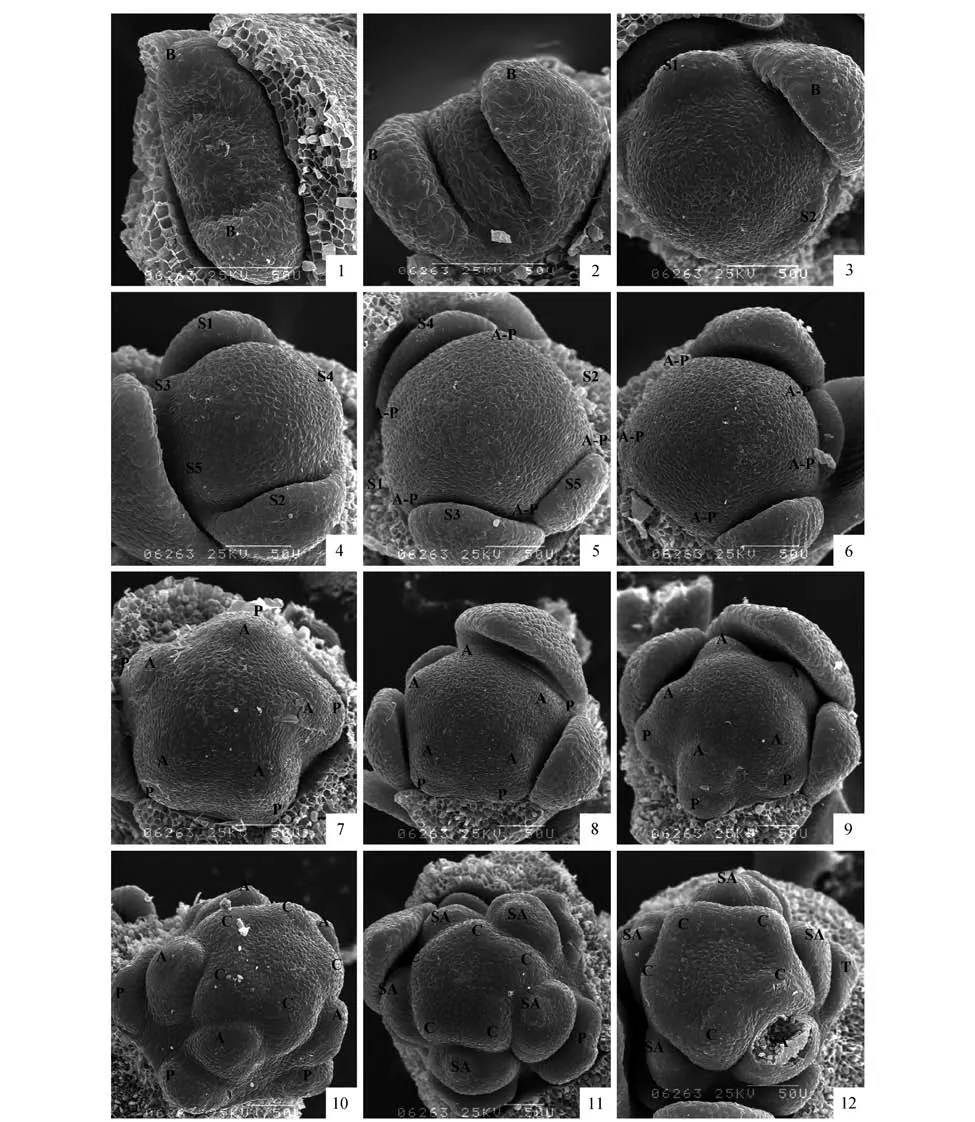
图版Ⅰ 扫描电镜下突脉金丝桃花器官的形态发生1~2. 花原基、苞片的发育及位置;3. 第1枚及第2枚萼片原基沿2/5圆周相继发生;4. 剩余3枚花萼原基的发生位置及形态;5~6. 雄蕊—花瓣共同原基开始发生;7. 雄蕊、花瓣原基近同时发生;8~10. 雄蕊、花瓣原基进一步发生,心皮原基开始发生;11~12. 次生雄蕊原基开始发生 B. 苞片;S. 萼片;P. 花瓣;A. 雄蕊;SA. 次生雄蕊;C. 心皮(字母后的数字代表发生顺序) 下同。PlateⅠ SEM photomicrographs of floral development in H.przewalskii1-2. Initiation of floral primordium and development and position of bract;3. Initiation position and shape of first pair of sepal primodia;4.Initiation position and shape of remaining sepal primodia;5-6. Initiation position and shape of androecium-petal common primordia;7. Initiation position and shape of petal primordia and stamen primordia;8-10. Further development of stamen and petal primordia and initiation of the carpel primordia;11-12. Initiation and development of the secondary stamen primordia B. Bract;S. Sepal;P. Petal;A. Androecium;SA. Secondary androecium;C.Carpe(lThe number indicates the initiation sequence of the floral organs after the letters) The same as below.
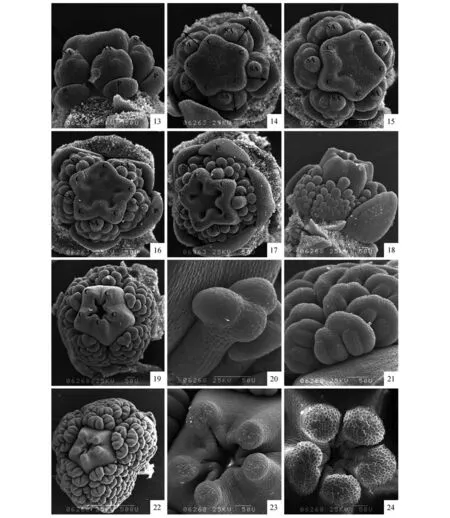
图版Ⅱ 扫描电镜下突脉金丝桃花器官的形态发生13. 次生雄蕊原基发生侧面观;14. 次生雄蕊原基的发生方式,箭头示次生雄蕊原基的离心发生;15. 心皮原基进一步发生,次生雄蕊原基数目增多;16~17. 心皮原基的进一步发生及雄蕊束形成;18. 雄蕊束形成侧面观;19~21. 次生雄蕊原基顶端出现人字形凹痕,花药开始形成,心皮对折加深;22~24. 分化出花柱和柱头,示柱头乳突细胞PlateⅡ SEM photomicrographs of floral development in H.przewalskii13.Lateral view of development of secondary androecia;14.Initiation position and mode of occurrence of the secondary androecium primordia. The arrow shows the centrifugation of the secondary androecium primordia;15. Further development of carpel primordia. Number of secondary androecium primordia increases;16-17. Further development of carpel primordia and fascicled stamen formation;18. Lateral view of fascicled stamen formation;19-21. Herringbone dents appear at the top of the stamen primordia,anther formation and the carpel folds deeper;22-24. Style and stigma differentiated,showing stigma mastoid cells
3 讨论
3.1 突脉金丝桃雄蕊束的发生与金丝桃属植物雄蕊束的起源
本研究发现突脉金丝桃的多数雄蕊是由次生雄蕊原基离心发生而来的,即5 枚雄蕊原基首先发生,待雄蕊原基隆起后,次生雄蕊原基由内向外依次发生,随着次生雄蕊原基的发育和数目的增多,可以观察到5 束雄蕊束逐步形成。此种发生方式还存在于金丝桃科及藤黄科的其他属中[15~16],与Leins[15]基于解剖学研究提出的金丝桃属植物雄蕊是离心发生的观点一致,也支持了雄蕊多数时,雄蕊呈离心发生的观点[17]。关于被子植物雄蕊束的起源前人进行了相关的研究,主要有三种观点:①聚合学说(Aggregation Theory):该学说认为多数的离生雄蕊是原始类型,离生雄蕊中有部分雄蕊不育而形成了间断的正常雄蕊簇生分区,然后由分区的雄蕊簇(Stamens cluster)聚合形成雄蕊束[18];②倍增学说(Dédoublement Theory):该学说认为固定基数的单轮雄蕊是原始的,而雄蕊束是由少数离生雄蕊经过分化加倍而形成的[19];③顶枝学说(Telome Theory):该学说认为雄蕊结构中的原始类型每组都是具有分支结构的,在进化的过程中每组分支结构的共同主干消失而形成了簇生的雄蕊束[20~22]。突脉金丝桃雄蕊束的发生过程表明,突脉金丝桃的雄蕊束起源于5 基数单轮雄蕊,支持了倍增学说的观点[19]。植物的花是不同进化阶段特征的混合体,花的发生和发育过程可以反映早期进化的轨迹,即研究个体发育可以为系统发育提供重要的线索[23]。Ronse Decraene 等[16]通过研究Harungana madagascariensis(金丝桃科)后提出广义藤黄科(包括金丝桃科)的雄蕊束可能起源于两轮雄蕊结构,雄蕊束起源于内轮雄蕊的分化加倍,而由于内轮雄蕊及花瓣的快速发育造成了外轮的雄蕊缺乏发育成熟的时间而退化为蜜腺。解剖学研究表明金丝桃属植物也存在着与Harungana 属相似的蜜腺结构[10,15],但Leins[15]认为金丝桃属植物的蜜腺是由花托二次发育而非退化雄蕊形成的。本研究在突脉金丝桃的花器官发生过程中也未观察到外轮对萼雄蕊原基的发生。因此本文认为金丝桃属植物的雄蕊束可能起源于单轮雄蕊而非二轮雄蕊。不过上述的结论仅仅基于金丝桃属的少数类群的研究得出,还缺乏整个金丝桃属花器官发生发育的证据,要真正解决金丝桃属雄蕊束的起源问题,还需进一步补充更多该属植物花器官发生的数据。
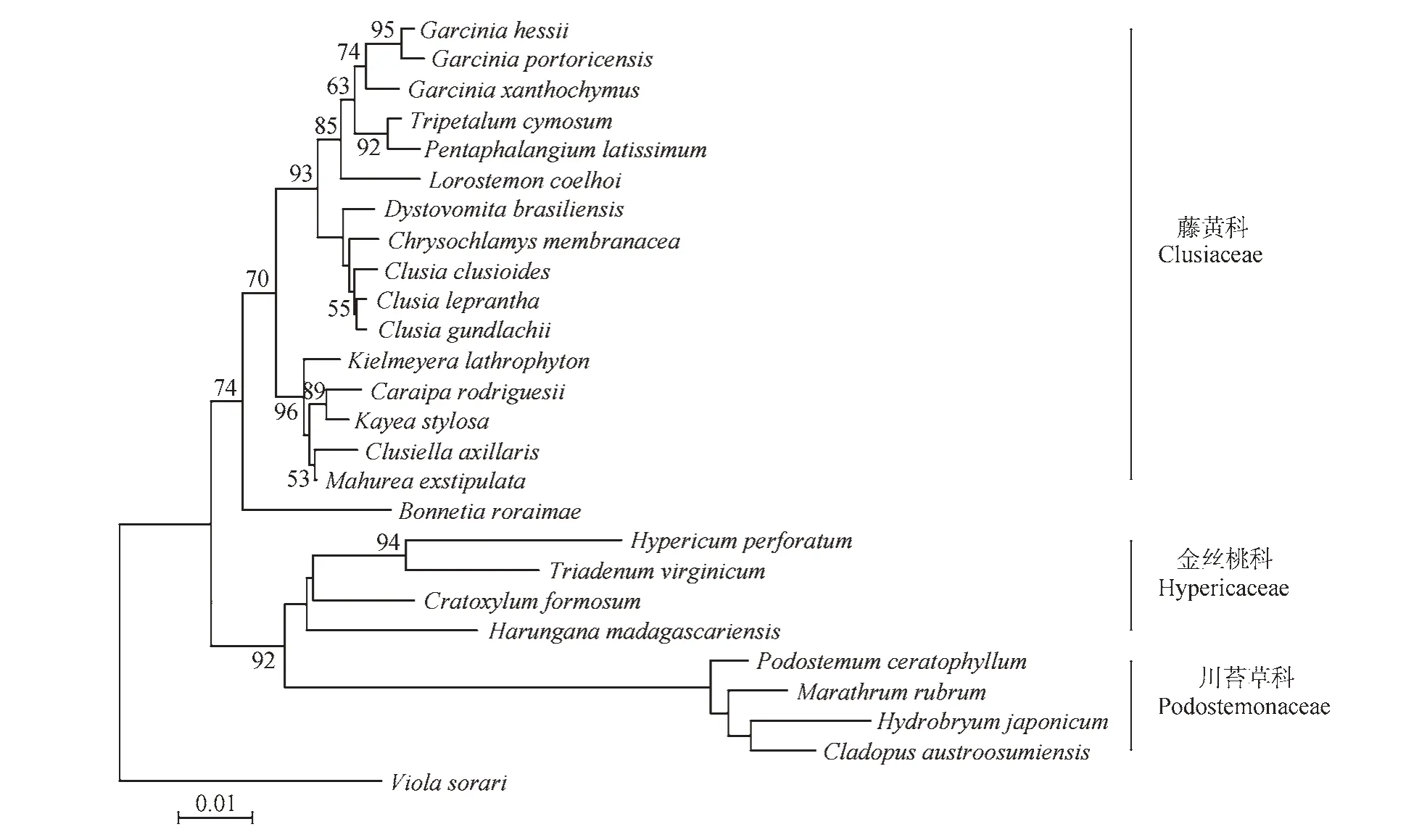
图1 基于rbcL序列构建的系统发育树[4]Fig.1 Phylogenetic tree constructed on the basis of complete rbcL gene sequences
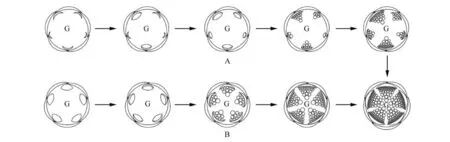
图2 花瓣及雄蕊束的发生方式简图A. 藤黄科;B. 金丝桃科;G. 心皮原基的位置Fig.2 Diagrammatic representation of the development of petal and fascicled stamensA.Clusiaceae;B.Hypericaceae;G.Position of carpal primordia
3.2 突脉金丝桃花器官发生的系统学意义
随着分子系统学的发展,有关金丝桃属分类学地位的研究出现了不同的观点。根据分子系统学的研究,APG Ⅲ系统将金丝桃属所属的金丝桃亚科从藤黄科中分出,独立为金丝桃科(见图1)[4~5]。成束的雄蕊是以上两科植物共同的花部特征之一[5]。本研究表明,突脉金丝桃的花瓣和雄蕊原基是由雄蕊-花瓣共同原基近同时分化发生的,其5 枚花瓣原基和雄蕊原基也为近同时发生(见图2)。此种发生方式也存在于金丝桃科的Harungana 属[16]。而Leins 等[12]研究发现藤黄科Garcinia volkensii 花瓣的发生过程与萼片类似,都是沿着2/5 的圆周相继发生,雄蕊原基在5 枚花瓣原基都隆起后才开始发生,发生方式与花瓣原基类似,但不同之处在于雄蕊原基的发生是在花瓣原基基部开始的(首先隆起的花瓣原基基部的雄蕊原基先开始发育),而花瓣原基却是在萼片原基之间的角隅处开始发生的(图2)。Hochwallner[24]在藤黄科书带木属(Clusia)中也发现了这种发生方式。虽然金丝桃科和藤黄科都存在成束雄蕊的结构,但二者花瓣原基及雄蕊原基的发生方式上却有着显著的不同,这也说明花成熟的结构只不过是发育过程中一个阶段的反映,形态学的研究应该树立动态的观点[11]。金丝桃科与藤黄科植物花瓣及雄蕊原基发生方式的显著不同,支持了APG Ⅲ系统将金丝桃亚科从藤黄科独立为金丝桃科的观点[5]。
