萘的微生物降解研究进展
2019-12-30刘小娜吴夏芫贾红华雍晓雨
刘小娜,李 彪,唐 晨,吴夏芫,周 俊,贾红华,雍晓雨
(南京工业大学 生物与制药工程学院,江苏 南京 211800)
多环芳烃(polycyclic aromatic hydrocarbons,PAHs)是一类具有高毒性和半挥发性的,且容易导致长期残留和生物蓄积的有机污染物,能够通过各种介质(生物体、大气、水体和土壤等)进行长距离迁移,在环境中分布极广,又难以降解,严重威胁自然环境与人类健康[1]。因此,近几十年来PAHs的污染治理始终都是全球性环境领域的研究热点之一。
萘(C10H8)是最典型的多环芳烃污染物,与其他多环芳烃物质一样具有极其严重的“三致”效应,即致癌性、致畸性及致突变性[2]。因此,萘被美国环境保护署列为须优先控制的16种多环芳烃类污染物之一,同时也成为多环芳烃降解的首要模型物及研究目标。
萘可以通过多种方法去除,如化学氧化、化学吸附、植物富集和生物降解等[3]。在上述几种方法中,生物降解由于具有操作简单、经济实用和不产生二次污染等优点,已逐渐成为近年来该领域的研究热点,也逐渐成为治理萘乃至多环芳烃污染的主要手段之一[4-5]。表1列举和比较了上述几种不同方法的原理与优缺点,其中萘的微生物降解通常又可以分为2种:一种是微生物,以有机萘为唯一碳源对其进行代谢;另一种是微生物将有机萘与其他有机质进行共代谢。

表1 萘的不同去除方法
1 萘降解菌株的分离
根据实际需求从不同生态环境中分离出萘降解菌株,对其降解效率及降解途径与机制的研究具有极其重要的科研价值和实际环保应用意义。表2为已经被分离、鉴定及广泛研究的常见萘降解菌株。目前分离到的萘降解菌株主要是细菌,如假单胞菌属(Pseudomonas)、红球菌属(Rhodococcus)、气单胞菌属(Aeromonas)、芽孢杆菌属(Bacillus)[10]等。
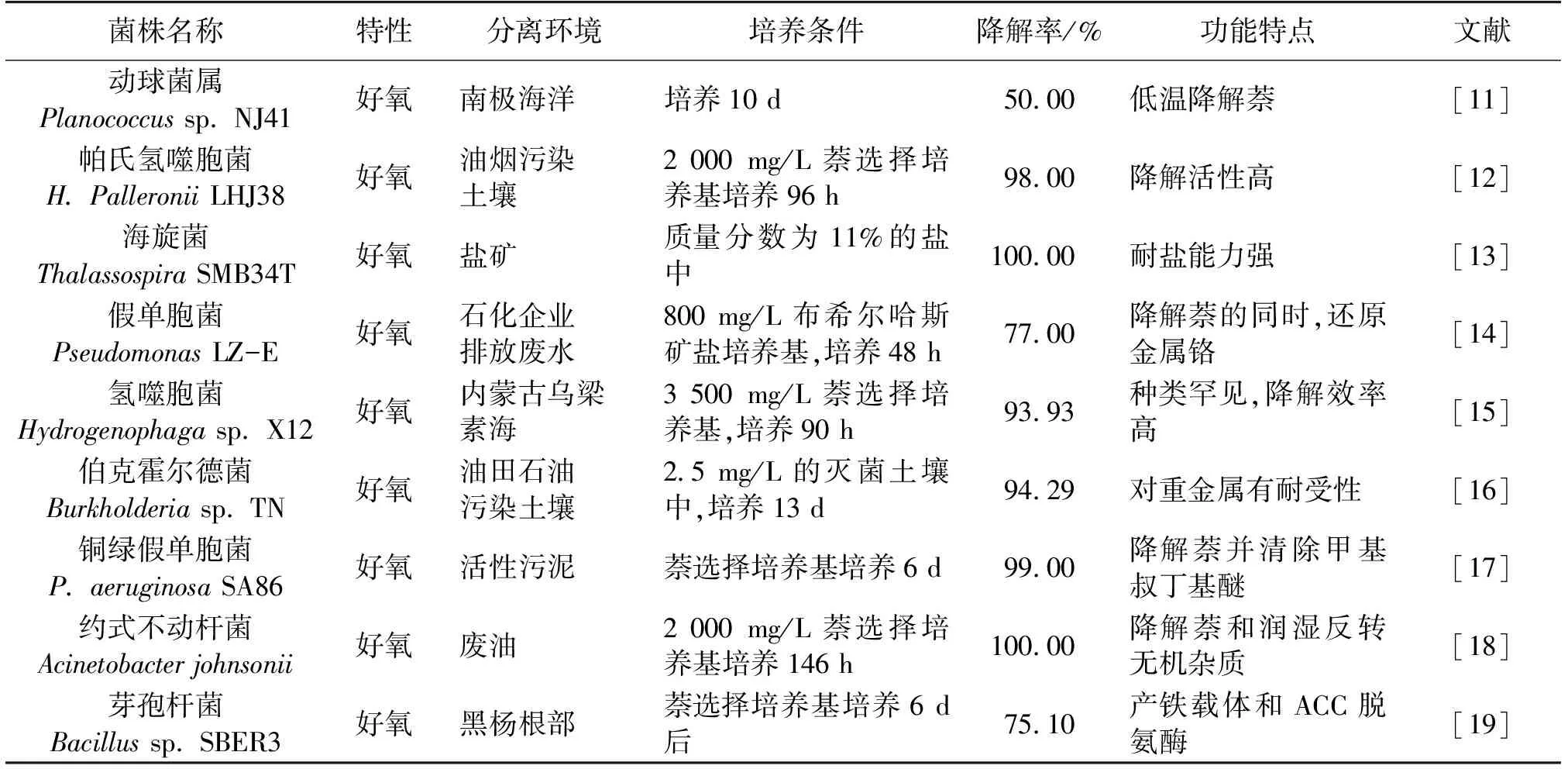
表2 不同环境中分离的萘降解菌株及其降解特征
1.1 自然环境中分离的萘降解菌株
在各种土壤、水体(包括湖泊、地下水)、植物根部甚至是极端条件等自然环境中广泛存在着萘降解菌株。2017年,孟建宇等[15]从来自内蒙古乌梁素海的样品中分离得到了氢噬胞菌X12,培养90 h后,该菌的萘降解率最高达到(93.93%)。王以斌等[11]以萘为唯一碳源从南极海洋水样中筛选出可低温降解萘(0~10 ℃)的动球菌属菌株Planococcussp. NJ41,该菌对萘的降解效率最高达到50%左右。此研究对于低温环境下的多环芳烃污染物降解及生物修复具有重要意义。Hadibarata等[20]从印尼热带雨林的土壤中也分离到一株同时具有降解萘和分泌木质素分解酶能力的白腐真菌(Pleurotuseryngii)。2014年,Kashir等[19]从美洲黑杨的根部分离筛选到一株具有萘降解能力的芽孢杆菌Bacillussp. SBER3,培养6 d后该菌的萘降解效率达75.1%。该菌在植物根部生长的过程中能够产铁载体和ACC脱氨酶,因此既可以促进植物生长又能降解萘。此外,土壤的肥沃程度也能够影响菌株降解萘的能力,Nakamura等[21]在含有丰富的有机物、生物碳及微生物的亚马逊中部原始土地上用萘丰富土壤进行微生物降解实验,发现含有生物碳的黑土地会提高萘的降解率。这可能是由于生物碳自身的多孔性,能够吸收大量微生物所需要的营养物质和水分及分布在周围的多环芳烃萘,为降解菌的生长提供了一个良好的生态环境[22]。
1.2 萘污染原位样品中分离的萘降解菌株
一些石油化工加工企业排放的废水及土壤中也分布着多环芳烃降解菌。宋昊等[12]从生活餐厨油烟污染的土壤中分离得到一株帕氏氢噬胞菌LHJ38,该菌在初始质量浓度为2 000 mg/L的萘选择培养基中培养96 h,萘降解率能达到98%;2011年,Plotnikova等[13]从俄罗斯彼尔姆地区的盐矿中分离到一株海旋菌ThalassospirapermensisSMB34T,能在浓度质量分数为11%的盐中与红球菌属(Rhodococcus)一起降解萘,降解率高达100%;Rahbari等[18]从Zayanderood河、Karon河以及伊斯法罕城市污水与活性淤泥中分离到一些能降解萘的假单胞菌株,其中铜绿假单胞菌P.aeruginosaSA86在降解萘的同时能有效清除与萘共存高毒污染物(甲基叔丁基醚),体现了该菌株的多功能性。张一博等[16]在油田石油污染土壤中分离到的一株高效萘降解菌伯克霍尔德菌Burkholderiasp.TN,该菌株能利用细胞壁上的活性基团富集Pb2+,适用于被萘和Pb2+双重胁迫的土壤修复。姜岩等[17]从废油中分离的一株约式不动杆菌在2 000 mg/L的萘选择培养基中能够100%降解萘,该菌株还具有润湿反转无机杂质的能力。2016年Huang等[14]从某石化企业排放废水中分离出一株假单胞菌LZ-E在降解萘的同时还可以还原重金属铬,是一株很好的复合污染修复菌株。一般来说,原位污染环境对菌株有长期的驯化,萘降解菌株能够成为该生态位中的优势菌株,因此降解萘的活性和效率也相对要高一些[23]。
2 不同因素对萘降解菌降解萘的影响
微生物对萘的降解受很多因素影响,这些因素通常都是对菌株的生长产生影响,例如外加营养物质、pH、温度和接种量等[24]。因此,针对萘降解菌株,对其培养的生长环境及条件进行研究与优化,可以优化目标菌株对萘的降解速率,从而使菌株的萘降解效率达到最大。
2.1 外加营养物质对生物降解萘的影响
目前,很多研究结果表明[25-27],添加适当的外源营养物质能够显著促进微生物对萘的降解率。贾燕等[26]发现向100 mg/L含萘水样中加入0.1 g NH4NO3、0.15 g KH2PO4、0.3 g K2HPO3、0.2 mL微量元素液,假单胞菌N7对萘的降解率提高23.65%。陈弘昊等[27]从辽宁盘锦红海滩地区翅碱蓬根系土壤及沿海滩涂土壤中分离出3株耐盐降解菌,分别为微杆菌属(Microbacteriumsp.S1)、恶臭假单胞菌(PseudomonasputidaY3)和刘志恒菌(Zhihengliuellsp. G12),当培养基中添加1.0 g/L葡萄糖时,3株菌对萘的降解率均达到最高值,且相对于对照分别提高了44.06% (S1)、70.56% (Y3) 和50.98% (G12)。孟建宇等[15]发现外加适量混合维生素对萘降解菌株生长及降解效率都有不同程度的促进作用。添加的营养盐、微量元素和生长因子等可以提供微生物生长的底物、代谢的能源物质、辅酶和调节细胞生长的因子等,显著提高菌株的有效生物量,从而从根本上提高菌株对萘的降解率[28]。
2.2 表面活性剂对生物降解萘的影响
萘的生物降解速率首先取决于其从固相向液相的迁移率。表面活性剂是具有固定亲水和亲油基团的两性分子,可以在溶液表面规则排列,防止油水之间的相互排斥。因此,表面活性剂因为自身结构的特殊性,可以降低有机萘分子的毛细管表面张力,提高疏水性物质在水相中的溶解度,增加其生物可利用性,进而提高萘的降解速率[29]。臧振远等[30]发现在添加质量浓度范围0.1~0.3 g/L的Tween-80能提高铜绿假单胞菌AEO-9对萘的降解速率。某些微生物在降解萘过程中能产生一定内源性的生物表面活性剂,其以糖脂形式(鼠李糖脂)存在,比外源表面活性剂的促进效果更优。这可能是由于自身产生的表面活性剂与微生物菌体的相适性较强,加快多环芳烃从固相到液相的迁移率,因此可以更快地促进萘的降解和微生物的生长[31]。
2.3 外加电子受体对生物降解萘的影响
呼吸是细胞将复杂的有机物逐渐分解成更小的化合物并最终生成CO2,释放出电子,同时驱动二磷酸腺苷(ADP)转化为三磷酸腺苷(ATP)的过程,释放的电子被电子受体捕获才能使这个过程持续进行[32]。在微生物好氧降解萘的过程中,O2作为氧化反应的终端电子受体(通过羟基化使萘开环)。因此,外通氧加大了电子受体的供应,使底物能够充分与优先电子受体氧接触,从而加快萘降解速率;另外适当的氧气含量也可以促进微生物对营养物质的摄取,加快微生物的生长[26]。贾燕等[26]研究发现把1 mL假胞单菌N7菌液接种到100 mg/L的萘选择培养基中,培养72 h,通氧量范围3.6~4.3 mg/L时,萘的降解率随着通氧量的增加不断升高,但当通氧量高于4.3 mg/L时,萘的降解率逐渐趋于平缓并保持平稳并态。因此,外源电子受体的供给量过高或过低都会影响降解率。另外,Kashir等[33]采用化学氧化法,在油田污染的地下水中加入1 750 kg的Na2S2O8等氧化剂后,SO42-浓度升高,萘降解可达到86%。证明外加氧化剂过硫化钠作为氧化反应的终端电子受体,可以加快萘的降解速率。
2.4 基本培养条件对微生物降解萘的影响
温度、pH、接种量和萘初始浓度等基本因素也显著影响微生物降解萘的效率。温度对于维持微生物体正常生命活动起关键作用。一方面,它能够影响微生物的生长速度及降解酶活性的高低来调节对外源物质的降解速度;另一方面,温度会通过影响底物的理化性质来影响底物运输速度[34]。雒晓芳等[35]发现枯草芽孢杆菌对萘的降解效果随着温度的升高得到提高,在培养温度为35 ℃时,降解效率最大,当培养温度继续升高时,降解率反而下降。因此,应该选择在最适温度下优化目标菌株对萘的降解效果。施氏假单胞菌YC-YH1在pH为7.0和8.0时降解率分别为100%和94.3%,在pH为5.0和9.0时的降解效率仅为19.3%和25.4%[36]。分析可能因为萘在降解过程中会产生少量有机酸并电离产生H+,导致溶液的pH值变小,过低的pH会引起微生物蛋白质、核酸等生物大分子电荷及原生质膜的电荷变化,影响辅酶和底物的解离程度,从而影响酶分子对底物分子的结合和催化;而pH过高又会对微生物吸收营养物质的能力和降解酶的分泌不利,最终影响微生物的生长速度及其代谢速率。同样,接种量对微生物降解萘也会产生很大影响。当接种量较少时,菌体生长迟缓期较长,降解速率也会相应较低;当菌体接种量较大时,会导致个别菌体吸收不到足够的碳源和能源,发生种内竞争抑制,从而使降解速率偏低。
3 萘的微生物降解途径
1964年Davies等[37]首次报道了微生物降解萘可能的途径,指出萘可以被微生物代谢生成多种降解中间产物,并最终被彻底降解成CO2和H2O。到目前为止,国内外报道微生物对萘的降解主要有3种途径,分别是水杨酸途径[38]、龙胆酸途径[39]和邻苯二甲酸途径[40](图1)。

Ⅰ—萘双加氧酶;Ⅱ—顺式萘二氢二醇脱氢酶;Ⅲ—1,2-二羟基萘双加氧酶;Ⅳ—2-羟基-2H-苯并吡喃-2-羧基异构酶;Ⅴ—顺-o-羟基苯脱萘丙酮酸水合-醛缩酶;Ⅵ—水杨醛脱氢酶;Ⅶ—水杨酸羟化酶;Ⅷ—邻苯二酚-2,3-双加氧酶;Ⅸ—邻苯二酚-1,2-双加氧酶;Ⅹ—水杨酸5-羟化酶;Ⅺ—1,2-双加氧酶;Ⅻ—异构酶/水解酶;ⅩⅢ—裂解酶;ⅩⅣ—双加氧酶/醛缩水合酶/脱氢酶)图1 萘的3种不同降解途径分析Fig.1 Three different degradation pathways of naphthalene
3.1 水杨酸途径
水杨酸途径是萘降解菌采用的最为常见的途径,多在假单胞菌属中发现。史延华等[24]以萘为唯一碳源分离出一株施氏假单胞菌YC-YH1,并且首次使用液相色谱-质谱联用(LC-MS)和化学需氧量(COD)的检测方法明确了施氏假单胞菌中萘降解产生的主要中间代谢产物,证明了其降解途径即为水杨酸途径(图1)。在由萘转化为水杨酸的上游代谢途径中,萘首先在萘双加氧酶的催化作用下开环形成顺-萘双氢二醇,然后被顺-萘双氢二醇脱氢酶催化形成l,2-二羟基萘,接着被l,2-二羟基萘双加氧酶催化转化为2-羟-2H-苯并吡喃-2-羧酸,并在2-羟-2H-苯并吡喃-2-羧基异构酶作用下再生成顺-o-羟基-苯亚甲基-丙酮酸,然后在顺-o-羟基苯脱萘丙酮酸水合-醛缩酶催化下形成水杨醛,最后水杨醛在水杨醛脱氢酶催化作用下生成水杨酸[41]。由于水杨醛脱氢酶的活性较低,因此该酶成为水杨酸途径中的关键酶和限速酶。萘降解下游途径中,水杨酸在水杨酸羟化酶催化下转化为邻苯二酚,邻苯二酚可以通过两种不同的代谢方式:一是在间位裂解酶邻苯二酚-2,3-双加氧酶的作用下开环形成己二烯半醛酸,最后形成乙醛和丙酮酸;二是在邻位裂解酶邻苯二酚-1,2-双加氧酶的催化作用下开环生成己二烯二酸,最后形成琥珀酸和乙酰辅酶A。上述2种不同路径所产生的产物最终进入三羧酸循环(TCA)后生成CO2和H2O[42]。
3.2 龙胆酸途径
龙胆酸途径也是萘降解的一条重要途径,其研究已有50多年的历史,截至目前已从最初细菌生化水平、酶学水平发展到近十几年的分子生物学水平揭示了微生物龙胆酸代谢途径和调控机制。青枯菌RalstoniaU2是龙胆酸降解途径的主要代表菌[43]。U2菌株能以萘作为唯一碳源生长,降解过程为萘在双加氧酶作用下降解成水杨酸(此过程与3.1章节水杨酸途径类似),随后水杨酸在水杨酸5-羟化酶[44]的催化作用下生成龙胆酸,并进入龙胆酸途径。龙胆酸在1,2-双加氧酶的催化作用下开环,生成中间产物顺丁烯二酸-单酰丙酮酸,然后在异构酶和水解酶的作用下形成丙酮酸和反丁烯二酸,接着进入TCA循环生成CO2和H2O[45](图1)。
3.3 邻苯二甲酸途径
在邻苯二甲酸途径中,萘首先在萘双加氧酶的催化下转化成顺式萘-二氢二醇,然后在顺式萘二氢二醇脱氢酶作用下生成1-2-二羟基萘并开环形成1-羟基-2-萘甲酸,然后以邻苯二甲酸途径代谢,随后在TCA循环中形成CO2和H2O[46](图1)。Abo-State等[47]以萘为唯一碳源筛选出一株博德特氏菌MAM-P22,用GC-MS分析萘降解中间产物,发现了邻苯二甲酸的存在。在Bacillusfusiformis[48]、Bacillusthermoleovorans中[49],也证实了邻苯二甲酸途径的存在。另外还有实验表明:羟基邻苯二甲酸是一种仅在邻苯二甲酸途径中产生的中间代谢产物(如铜绿假单胞菌),总体而言,邻苯二甲酸微生物萘降解途径仍然在继续解析中。
4 萘的关键降解酶及其编码基因
截至目前,对萘降解菌降解途径中的关键酶及其编码基因的研究已经非常广泛。其中萘双加氧酶是催化萘降解途径第1步反应所需要的关键酶,且是3个途径中的必要酶。该酶由4个亚基构成,分别为萘双加氧酶铁氧还蛋白、萘双加氧酶还原酶、萘双加氧酶Fe-S蛋白大亚基以及萘双加氧酶Fe-S蛋白小亚基[50]。萘双加氧酶由nahAaAbAcAd基因编码,有研究表明所有的萘降解菌株都含nahAc基因[51]。萘趋化蛋白基因nahY同样发挥着重要的作用,它可以使细菌向萘及其代谢中间产物泳动,促进其对萘的降解,从而加快降解速率[52]。在萘降解的水杨酸途径中,催化邻苯二酚间位裂解的酶基因是邻苯二酚2,3-双加氧酶基因nahH[53],催化邻苯二酚邻位裂解的酶基因是邻苯二酚1,2-双加氧酶基因catA[54]。水杨醛脱氢酶由nahF编码形成,其催化水杨醛脱氢生成重要中间产物水杨酸,是萘降解上游途径操纵子的关键基因。nahG是萘降解途径下游操纵子的第1个基因,它编码的水杨酸羟化酶催化水杨酸羟化生成邻苯二酚。2006年,李永君等[51]从污水处理厂的活性污泥和石油工业废水中各分离出24株萘降解菌,分别提取这些菌株的总DNA,然后与各种萘降解基因杂交。发现来自工业废水的萘降解菌株均含有萘双加氧酶的铁硫蛋白大亚基基因nahAc、水杨醛脱氢酶基因nahF及其重复基因nahV、水杨酸羟化酶基因nahG及其重复基因nahU、儿茶酚2,3-双加氧酶基因nahH、儿茶酚1,2-双加氧酶基因catA以及萘趋化蛋白基因nahY。而来自活性污泥的萘降解菌株只含有nahAc、nahF、nahG和catA,不含有nahY、nahV、nahU和nahH。来自石油废水的萘降解菌株降解活力和稳定性明显比污水中的降解菌株强。另外具有重复降解基因nahV和nahU的细菌对环境具有更好的适应能力。Bosch等[55]发现在若干P.stutzeri菌株中都存在水杨酸羟化酶基因nahG及重复基因nahW,因此推测所有萘降解菌中可能都存在该重复基因。
2017年马鹏等[56]用萘选择培养基对淮东石油污染的土壤中萘降解菌进行富集培养,采用萘双加酶基因通用引物对该目标基因片段进行扩增、测序及比对分析,验证了筛选出来的菌株是萘降解菌株。以上研究表明:可以选择萘降解途径中的关键酶基因对菌株进行PCR扩增或DNA杂交来初步判断、筛选萘降解菌株,并可以通过对关键代表性的功能基因进行序列比对分析初步推断该目标菌株的萘降解途径,表3为关键酶基因引物序列。

表3 萘微生物降解途径中关键基因的引物序列
5 萘微生物降解的实际应用
萘作为优先控制的环境污染物,其微生物降解技术在环境治理及资源利用等方面的实际应用尤为重要。李春霞[59]从吉林省松原油田污染的土壤中分离筛选出一株降解萘的绿脓假单胞菌(P.aeruginosa),将该菌株应用在实际污染土壤样品中,研究发现该菌株对萘的降解率为60.4%,这一研究对降低或消除环境中萘污染的生物修复起了一定的推动作用。2015年,廉景燕等[60]从大庆油田石油污染土样中分离得到了一株高效降解萘的假单胞菌属(Pseudomonassp.)DQ-1,并通过室内模拟修复实验对该菌株的土壤修复能力进行了研究。研究发现:加入DQ-1菌液明显提高了土壤中萘的降解速率,且菌株对重金属有一定的耐受性,在萘和重金属复合污染的土壤修复中有一定的实用价值。总体而言,萘微生物降解的研究目前主要局限于实验室中,亟须要将该研究转化为有实际应用价值的技术。
6 萘微生物降解研究存在的问题及展望
至今为止,科学家们对萘微生物降解的研究内容已十分广泛。已从萘降解菌株的筛选、分离、培养条件及降解性能的优化和降解途径的挖掘,逐步转移到萘降解关键基因及酶功能解析、降解调控因子及其分子遗传机制的理论研究中。
在笔者看来,该领域的研究还需要借助现代高通量测序及生物信息学分析,在基因组、转录组及代谢组学水平将菌株生长、代谢和降解等功能途径有机整合,进一步从合成生物学角度建立降解功能元器件模块、构建萘降解底盘高效微生物。同时,目前对于厌氧条件下萘降解的研究相对较少。厌氧萘降解菌株广泛分布在于植物根际、地下沉积物、海洋和极地土壤中,它们凭借多样的类型、高效的代谢能力与多功能的生理特性在环境保护及资源利用等方面表现出巨大的应用潜力。
因此,应该加大对萘微生物厌氧降解的研究,以达到对一些被污染的特殊生境的修复。最后,应当加快萘污染场地微生物修复的工程化应用研究,在处理实际问题的过程中解决微生物修复技术所面临的菌株存活难、修复周期长和稳定性差等问题,提高萘微生物降解的实际应用效果与价值。
