不同光照条件下浒苔与三种赤潮微藻的竞争
2019-08-28高颖超谭丽菊王江涛
高颖超,杨 雪,林 锟,谭丽菊,王江涛
不同光照条件下浒苔与三种赤潮微藻的竞争
高颖超,杨 雪,林 锟,谭丽菊,王江涛*
(中国海洋大学化学化工学院,山东 青岛 266100)
针对苏北浅滩水体浊度高、营养盐含量大、且被大多数学者认为是南黄海绿潮起源的实际情况,本文通过室内培养实验,研究了不同光照条件(0~4500lx)下浒苔与中肋骨条藻、东海原甲藻和新月菱形藻对营养盐的竞争作用;并与苏北浅滩实际水体光照相结合,评估了浒苔与3种微藻在不同深度下的生理状态.浒苔与微藻的竞争,是决定暴发何种藻华的关键因素之一.结果表明,与3种微藻相比,浒苔对光照的适应性更强,最大日均相对增长率为11.91%/d.微藻与浒苔共培养时,生长情况明显劣于单独培养, 中肋骨条藻、东海原甲藻和新月菱形藻的生长抑制率分别为30%~46%,3%~44%,25%~ 41%,并且不同微藻对竞争作用的响应不同.营养盐监测数据表明,对有限营养盐的争夺是浒苔和微藻竞争作用的主要机制,研究结果为苏北浅滩浒苔绿潮的暴发机制提供了依据.
浒苔;微藻;光照强度;营养盐;竞争作用
在过去的十几年中,浒苔绿潮在我国南黄海呈现连续性和常态化的暴发趋势,给我国沿海城市的居民生活、水产养殖业、旅游业等带来诸多负面影响.研究表明,同一海域内大型海藻生物量与浮游微藻生物量往往呈负相关关系[1],通常是由化感作用以及对生源要素的竞争作用导致[2-3].在藻华暴发初期,大型海藻与浮游微藻的竞争作用决定了藻华的优势种,因此有必要分析暴发初期浒苔与微藻间的竞争,以期为绿潮灾害的预防和治理工作提供借鉴.大量研究证实,光照、温度和营养盐等环境因素在浒苔萌发阶段起着决定性的作用[4].
目前,对浒苔单独培养体系的研究报道较多[5-7].结果表明,高营养盐和低盐度能够协同促进缘管浒苔的光合作用[8];浒苔对高温和高光的耐受性较好,在15~25℃、200~600μmol/(m2·s)条件下其比生长速率是扁浒苔、曲浒苔、缘管浒苔的1.5~3.5倍[9];浒苔可以直接吸收多种形态氮营养盐,吸收速率依次为NO3−> NH4+> CO(NH2)2> C2H5NO2和C3H7NO2[10];重金属如Cu、Zn的添加对浒苔吸收硝酸盐具有低促高抑作用[11].
在实际海域中,往往是浮游微藻与大型藻类共同存在,目前关于二者共存时对营养盐竞争的研究较少.且苏北浅滩水体含沙量大[12],浑浊度高,光照强度与其他海区明显不同,本文调查结果也表明,大部分海域在0.5m深度处透明度就几乎为零.在此特殊水体条件下,浒苔和微藻对光的适应性及其对营养盐竞争能力的差异,就成为暴发何种藻华的关键因素.本文在实验室模拟条件下,研究了浒苔和3种典型赤潮微藻在不同光照下的竞争作用,同时将室内培养结果与苏北浅滩实际水体光照相结合,对浒苔与3种微藻在不同深度下的生理状态进行了评估,分析了环境因素对苏北浅滩绿潮暴发机制的影响,为浒苔防控和治理提供理论基础.
1 材料与方法
1.1 实验材料
实验用中肋骨条藻()、东海原甲藻()、新月菱形藻()均取自中国海洋大学海洋污染生态化学实验室,用藻类培养基f/2培养基[13]培养至指数生长期待用.实验用浒苔()于2017年5月采自苏北浅滩,采集后用灭菌海水反复清洗3~4次,以去除泥沙和其他附着生物,镜检确认无杂藻后,挑取生长状态良好的藻体备用.实验用海水采集自苏北浅滩,用孔径为0.45μm的醋酸纤维滤膜过滤,经120℃高温灭菌20min,自然冷却后使用.海水本底NO3--N和PO43--P浓度分别为49.51μmol/L和1.89μmol/L,根据测得的营养盐浓度重新添加营养盐达到f/2培养基的要求,同时添加1mg/L GeO2以消除硅藻的影响[14].
1.2 实验仪器
ZDS-10W-2D照度计(上海市嘉定学联仪表厂),GXZ-500B恒温光照培养箱(宁波市江南仪器厂),TechniconTMAA-Ⅲ型营养盐自动分析仪(德国BRAN+LUEBBE公司),DM4000B电子显微镜(德国徕卡公司),AL204电子分析天平(瑞士METTLER TOLEDO公司),LDZX-50FBS立式压力蒸汽灭菌器(上海中安医疗器械厂).
1.3 实验设计
实验设置5组光照梯度,侧面进光,通过在培养瓶外包覆不同颜色的遮光布来达到遮光目的.测定培养液中的光强,使光照强度分别为0,900,1800,3600, 4500lx,每组重复测定10次.同时于2017年5月赴江苏盐城实地调查,测量浒苔暴发海域不同水深的光照强度,并推算实验室光照条件对应的平均水深.表1中列出实验所用遮光布的透光率情况.
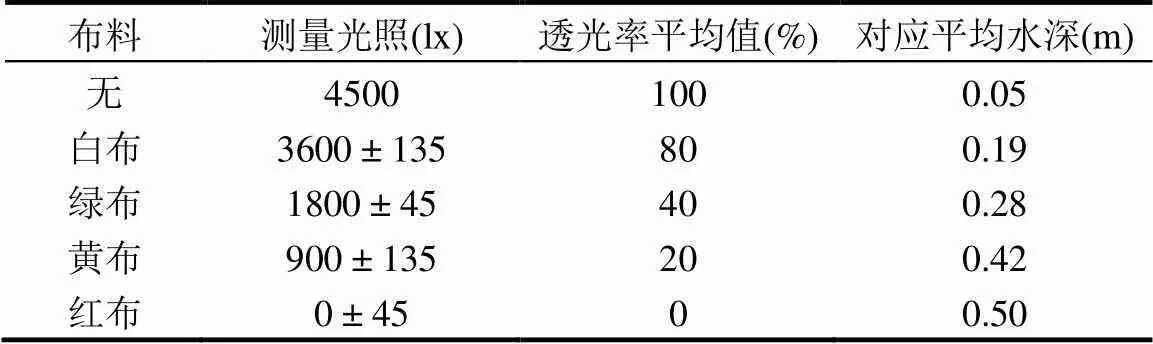
表1 不同遮光布的透光率
实验设置7组培养体系,如图1所示,分别为浒苔单独培养(UPC);中肋骨条藻单独培养(PC-S);东海原甲藻单独培养(PC-P);新月菱形藻单独培养(PC-N);浒苔-中肋骨条藻共培养(MC-S);浒苔-东海原甲藻共培养(MC-P);浒苔-新月菱形藻共培养(MC-N).

光照强度0lx900lx1800lx3600lx4500lx
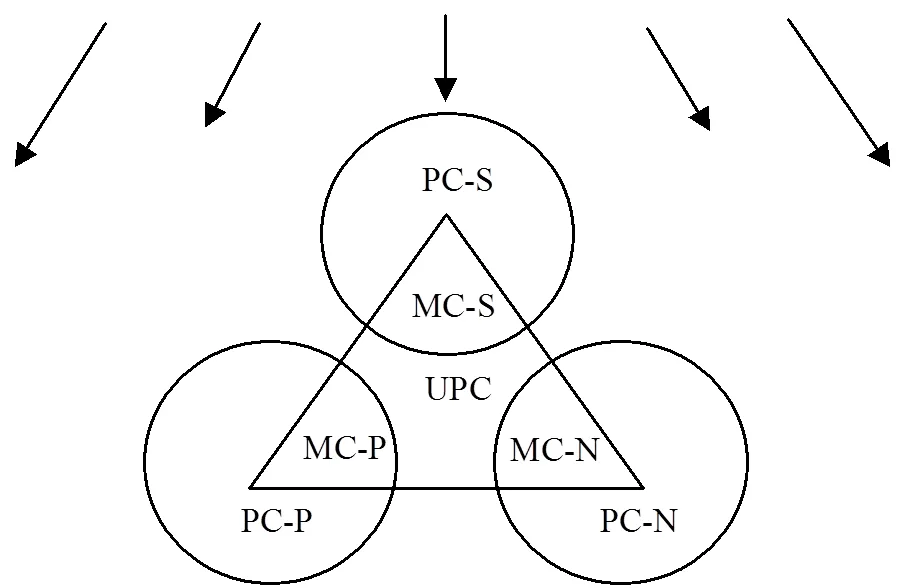
图1 培养体系示意 Fig.1 Incubation series
1.4 实验方法
取生长状况良好且处于指数生长期的微藻和/或新鲜的浒苔藻体接种于150mL f/2培养基中,在不同的光照强度(图1)下培养.培养温度为(20 ± 0.1)℃,光照周期12L:12D.浒苔初始接种量为1.0gFW/L,记录微藻初始接种藻密度,每组设计3个重复,实验共进行7d.每天定时摇动培养瓶3次,以防微藻附壁生长并保持溶解气体含量.
1.5 生物量测定
自接种当天起,每24h用镊子取出浒苔藻体置于吸水纸上吸干表面水分,放在干燥的培养皿中称量藻体湿重,按公式(1)计算浒苔的相对增长率[5]:
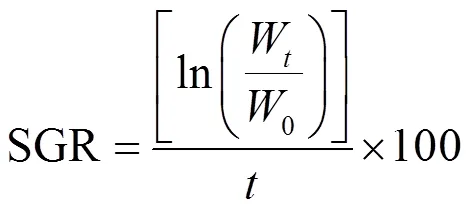
式中:0为浒苔初始鲜重,g;W为实验时刻浒苔鲜重,g;为培养时间,d.
同时从培养瓶中取藻液2mL并用Lugol’s试剂固定,采用浮游生物计数框在显微镜下计数藻细胞密度,每个样品计数4次.生长抑制率计算方法[15]为:
= (1 –0)×100% (2)
式中:为处理组生物量;0为对照组生物量.
1.6 营养盐测定
培养液营养盐浓度采用营养盐自动分析仪进行测定,并根据公式(3)计算营养盐吸收速率[16]:
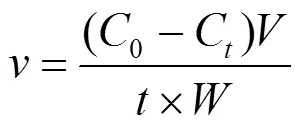
式中:表示吸收速率,μmol/(g·h);0和C分别是实验开始时与实验时刻的营养盐浓度,μmol/L;为取样时的培养液体积,L;为浒苔湿重,g.
1.7 数据处理
实验数据使用SPSS 19(IBM Statistical Package for the Social Sciences Inc.)进行统计分析,比较不同培养体系中浒苔和微藻相对增长率的差异(one- way ANOVA),探讨光照强度对浒苔和微藻生长及竞争的影响.
2 实验结果
2.1 浒苔生物量的变化
在其他培养条件相同时,不同光照条件下浒苔生物量变化及日均相对增长率见图2.从图中可看出,在UPC培养体系中,光照越强,浒苔日均相对增长率越大,但在3600~4500lx的范围内光照强度对浒苔生长率的影响差别不大(>0.05).在单独培养前期(0~2d),浒苔湿重呈近似线性增长,其中3600lx时浒苔最大相对增长率为10.50%/d,4500lx时为11.91%/d,随后浒苔生长进入平稳期(2~7d),可能是受到了营养盐的限制作用.通过对培养液中营养盐含量测定发现,光照强度对浒苔吸收营养盐有一定的影响,光照强度为4500lx时营养盐消耗最快,0lx时消耗最慢,综合图2(a~d)分析可以看出浒苔生物量与营养盐消耗速率相关.NO3-N吸收速率在培养前3d快速增加,然后以1.0μmol/(g·h)左右的速率稳定.在整个培养实验周期内,PO43--P不断被消耗,吸收速率维持在0.10μmol/(g·h)左右(3600lx),适宜光照(3600,4500lx)条件下PO43--P几乎被消耗殆尽,因此实验末期浒苔生物量下降可能是由于营养盐耗尽导致,特别是受到磷的限制作用.
在其他实验条件均一致时,相比于微藻(图3、4、5),浒苔更能适应低光照条件,但在光照强度为0lx时,浒苔也不能正常生长,日均相对增长率为-0.21%/d,由此结合现场条件进行推断,当浒苔下沉至水深0.50m以下时可能无法正常生长.
浒苔与微藻共培养时,生长率有所降低,结合图2(e)发现,浒苔与不同微藻共培养时产生的响应不同.与中肋骨条藻共培养时,浒苔生长并没有受到很大影响,日均相对增长率下降0.50%左右.因此当浒苔暴发海域有中肋骨条藻共存时,浒苔在0.50m深度内能正常生长.与东海原甲藻共培养时,除了4500lx光强下浒苔有一定增长外,其余光照条件下浒苔基本不生长,最大相对增长率出现在4500lx光强下的前24h,为3.48%/d,因此当浒苔暴发海域出现东海原甲藻大量繁殖时,浒苔可能仅在0.05m深度内能正常生长.与新月菱形藻共培养时,不同光照条件下浒苔生长均受到抑制,呈现负增长,最大抑制率同样出现在高光强4500lx下,为44.70%.同时新月菱形藻生长也受到一定抑制,可能是由于高光胁迫导致藻细胞破裂,释放某种胞内物质抑制浒苔生长,可以观察到部分藻体发黄甚至死亡,由此推断当浒苔暴发海域有新月菱形藻共存时,浒苔在不同深度下的生长均可能受到抑制.
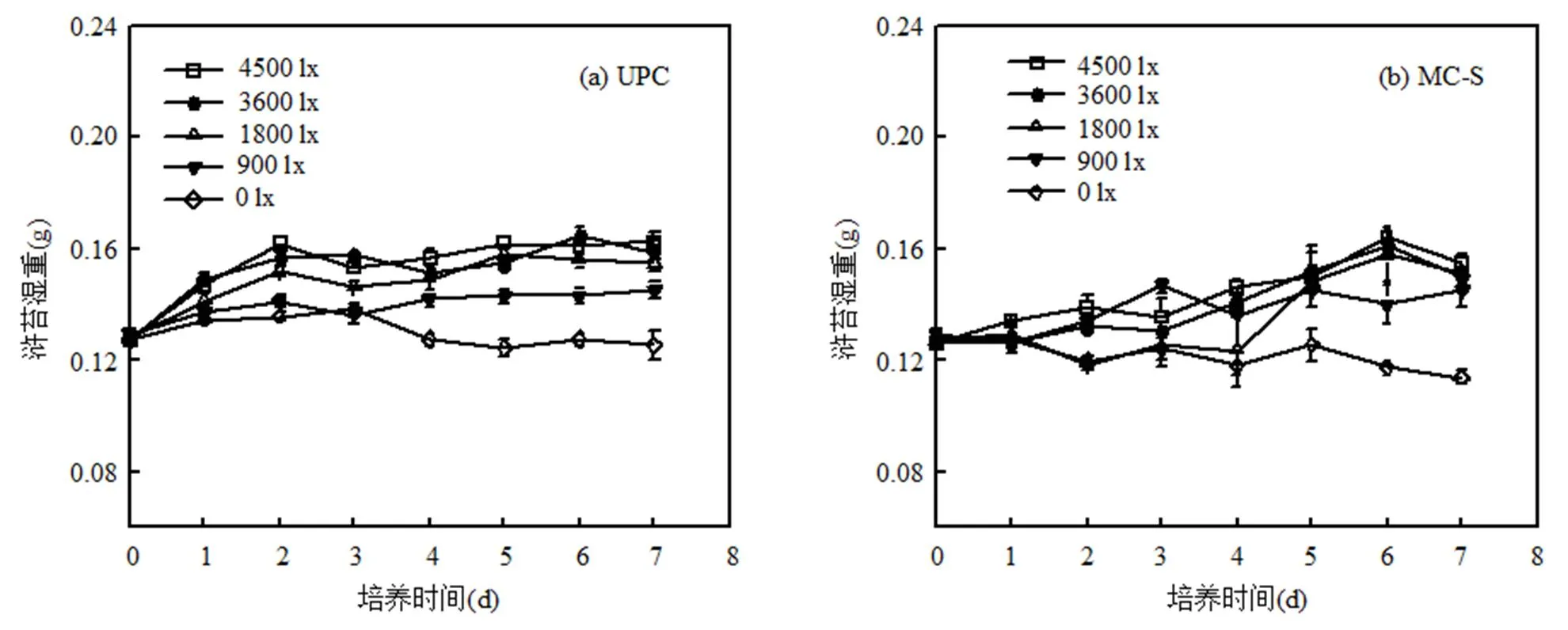
图中a、b、c、d代表差异显著性,不同字母表示存在显著性差异,<0.05
2.2 微藻生物量的变化
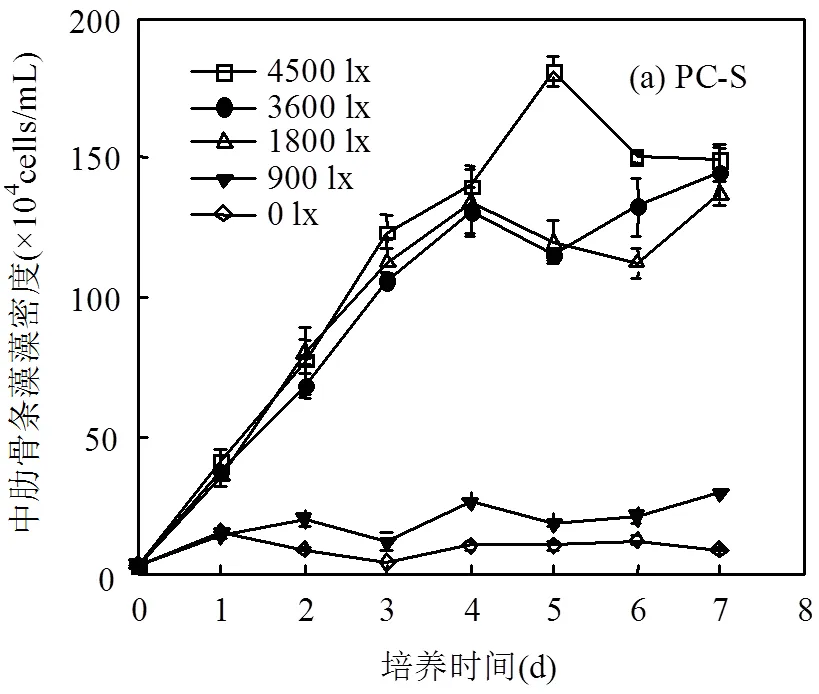
2.2.1 中肋骨条藻藻密度的变化及日均相对增长率 图3(a)为单独培养条件下中肋骨条藻藻密度变化曲线,由图可见中肋骨条藻对光照的适应范围不如浒苔,在低光照强度(0和900lx)下,中肋骨条藻几乎不能正常生长.其余光照条件下培养前4d,中肋骨条藻均以指数速度迅速增长,最大相对增长率可达到100%~110%/d;第5d藻密度开始出现不同程度下降,可能是受营养盐(尤其是磷、硅)的限制作用.
图3(b)为共培养体系中肋骨条藻藻密度变化曲线,由图可见,中肋骨条藻在前2d生长停滞,在第3、4d开始快速生长,随后进入平稳期.由图3(c)可知共培养体系中中肋骨条藻的日均相对增长率劣于单独培养体系,生长抑制率为30%~46%,说明在共培养体系中中肋骨条藻生长受到了一定抑制.通过对MC-S培养体系中营养盐测定发现,培养前期培养液中营养盐浓度急剧下降,培养1d后培养液中硅浓度仅为加富硅浓度的一半,前24h硅的吸收速率可达到1.7μmol/(g·h)左右,浒苔并不能利用硅,说明中肋骨条藻生长吸收利用硅极快.在培养的第5d,培养液中的P、Si均已耗尽,结合图3(a,b)可以看出此时中肋骨条藻的生长受到了一定抑制作用, NO3-N浓度也比UPC培养体系中要低,因此培养体系中浒苔与中肋骨条藻的营养竞争可能是导致培养后期中肋骨条藻密度不再上升的原因,但也不排除化感物质的干扰[17-18].推断在现场条件下,中肋骨条藻可能在0.42m深度左右生长即受到抑制,并且由于浒苔的遮光或其他作用,在小于0.42m深度时即可能无法正常生长.
2.2.2 东海原甲藻藻密度的变化及日均相对增长率 由图4可见,在光照强度较低(0,900lx)时,东海原甲藻生长状况也明显较差.在单独培养体系中,当光照适宜时东海原甲藻藻密度呈指数递增趋势,日均相对增长率为30%/d左右.
图中a、b代表差异显著性,不同字母表示存在显著性差异,<0.05
结合图4(a,b)可看出,共培养体系中东海原甲藻对浒苔的响应十分敏感,培养4d后才开始生长.从抑制率数值上来看,共培养显著抑制了东海原甲藻的生长,抑制率为3%~44%,镜检发现藻细胞出现严重受损.培养液中营养盐的测定结果显示,NO3--N和PO43--P浓度变化趋势基本相似,从培养的第2d开始,营养盐吸收速率升高,培养中期(2~4d)NO3--N吸收速率为2.0μmol/(g·h), PO43--P吸收速率为0.21μmol/(g·h);培养后期(5~7d) NO3--N吸收速率为7.3μmol/(g·h),PO43--P吸收速率为0.42μmol/(g·h),可能是由于后期东海原甲藻藻密度上升导致消耗增加.从营养盐的变化情况来看,浒苔与东海原甲藻共培养时,营养盐始终是充足的,因此东海原甲藻生长缓慢可能不是因为营养竞争而是受到浒苔分泌的化感物质的影响,并且此化感作用可能与浒苔的生长增殖有关[17-19].在现场环境下,东海原甲藻可能在0.28m深度内能正常生长,当与浒苔共存时,其生长将受到抑制.
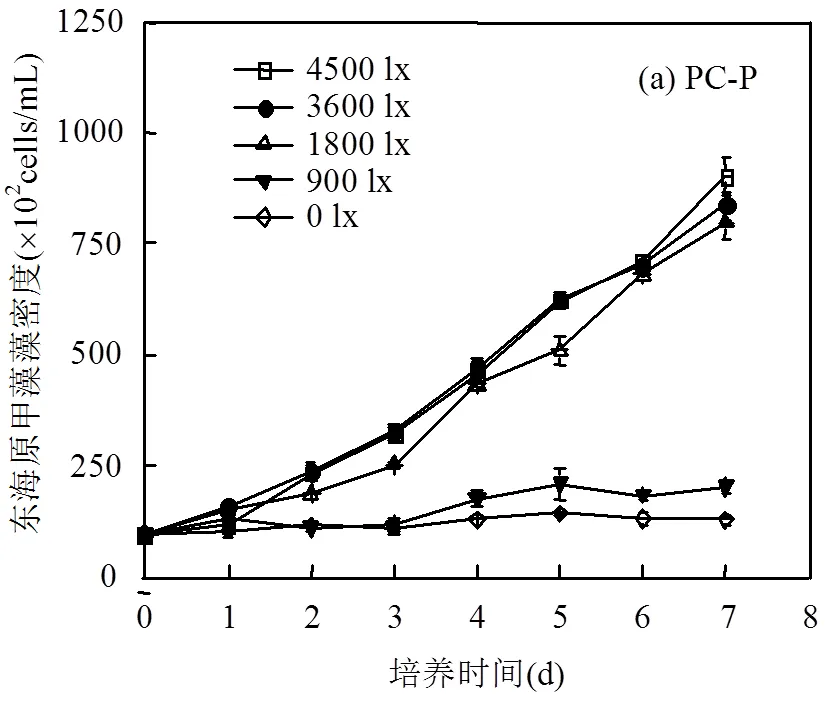
图中a、b代表差异显著性,不同字母表示存在显著性差异,<0.05
图中a、b代表差异显著性,不同字母表示存在显著性差异,<0.05
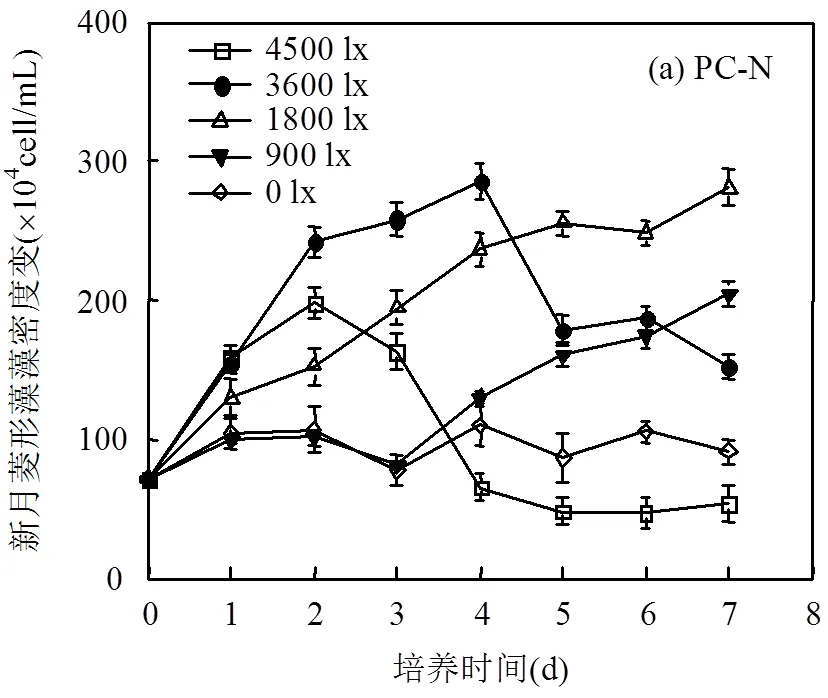
2.2.3 新月菱形藻藻密度变化及日均相对增长率 新月菱形藻对光照强度的适应范围明显区别于其他2种微藻,由图5可见,光照强度为1800lx时新月菱形藻生长最好,日均相对增长率达到19%/d,高光强(4500lx)或低光强(0lx)下新月菱形藻生长均受到一定程度的抑制.4500lx条件下新月菱形藻生长受到的抑制作用最明显,日均相对增长率甚至不如低光强(0lx)时,可能是由于高光胁迫导致新月菱形藻细胞受损,藻密度迅速下降.
在光照强度适宜(900~3600lx)时,共培养体系中新月菱形藻生长情况明显劣于单独培养,生长抑制率为25%~41%.共培养体系中,新月菱形藻能很快适应新环境,培养前期生长未出现停滞,并且培养第2d藻细胞数达到峰值;培养中后期,新月菱形藻生长受到抑制.通过对培养液中营养盐测定发现,营养盐吸收速率基本可分成2个阶段,即前期快速吸收阶段和后期平稳吸收阶段,与浒苔和新月菱形藻的生物量变化相关.在培养前24h, NO3--N吸收速率为2.1~ 7.3μmol/(g·h), PO43--P吸收速率为0.40~0.58μmol/ (g·h), SiO32--Si吸收速率也能达到2.0μmol/(g·h)左右.培养后期营养盐含量相对充足,但新月菱形藻生长受到持续抑制,并且抑制作用随着培养时间不断加剧,实验结束后培养液中营养盐虽然没有全部耗尽,但也被大量吸收,因此考虑新月菱形藻在培养中后期受到的抑制可能是营养竞争和浒苔分泌的化感物质的影响耦合所致[20-21].结合现场情况来看,新月菱形藻仅能在0.28~0.42m左右深度处存活,过深或过浅都不能正常生长,与浒苔共存时,其生长也将受到抑制.
3 讨论
研究资料证实,大型藻类与海域中微型浮游植物的相互作用在海洋环境中是普遍存在的,作用机制包括资源竞争和释放化感物质,甚至是两种作用相互耦合的结果[22-23].藻类之间的竞争除了营养盐以外,还受温度、光强、pH值等因素的影响.光照是海洋浮游植物生长的能量来源基础,是其能够进行光合作用的必要条件.在一定范围内,随着光照强度的增加,浮游植物的生长速率也会增加,但光照强度过高或过低都会限制浮游植物的生长.
苏北海区水深较浅,风浪引起的沉积物再悬浮等因素可减弱水体光照强度[12].本文结果显示,在苏北浅滩的特殊水体光照条件下,浒苔对光照强度的适应性及对有限营养盐的竞争能力要优于微藻,同时在绿潮暴发期间,浒苔大规模繁殖漂浮在海面上,大量耗氧遮光,导致其他海洋微藻不能正常进行光合作用,这可能也是使海洋微藻生物量降低的原因之一.本研究在不同光照水平下将浒苔与微藻共培养,可验证苏北浅滩特殊水体光照条件下浒苔与微藻的相互作用.
此外,光照强度和光照周期都会影响浮游植物对营养盐的吸收利用,尤其是氮营养盐[24-25];光照强度影响营养盐吸收形态及速率,光照周期则会影响相关酶的活性及合成.硝氮是浒苔主要吸收利用的营养盐;氨氮的吸收受光照影响较小,即使是黑暗条件下也可以被浒苔吸收利用[26],因此本实验不予讨论.近年来研究发现,磷限制是全球海洋存在的一种普遍现象[27],已有研究表明,黄海海域自浒苔暴发后海水中PO43--P水平一直处于限制状态[28].庞秋婷等[7]在围隔实验培养中也发现浒苔对PO43--P具有极强的吸收能力,PO43--P浓度偏低时会限制浒苔的生长.因此,营养盐限制对浒苔生长的影响作用十分重要,本研究中,浒苔相对增长率的结果与营养盐利用程度显著相关(<0.05).
本文结果显示,不同微藻与浒苔竞争作用机制不同,浒苔与中肋骨条藻共培养时,中肋骨条藻生长受到一定抑制,浒苔生长基本不受影响,考虑为营养盐竞争导致,但也不排除化感物质的干扰;浒苔与东海原甲藻共培养时,东海原甲藻相对增长率降低甚至出现死亡,浒苔生长也十分缓慢,而营养盐始终充足,考虑是受化感物质的影响;浒苔与新月菱形藻共培养时,两者生长均受到显著抑制,并且抑制作用与光照强度有关,培养液中营养盐也被大量吸收,考虑可能是营养竞争和化感作用耦合的结果.在营养盐充足情况下,相对于东海原甲藻,中肋骨条藻能更好的利用营养盐来进行快速增殖,这与其他人的研究一致[29-31].本实验中浒苔相对增长率出现负值,可能与浒苔的生长阶段有关,在以后的研究中将尽可能选用采自苏北浅滩的浒苔幼苗来进行实验.浒苔与3种微藻在不同深度下的生理状态是基于室内培养实验结果进行的估计,为现场调查研究提供了一定指导意义.另外,共培养体系中浒苔占优势是否与化感物质有关,还需通过今后研究浒苔生长过程中的分泌物质来进行验证.
4 结论
光照强度是限制藻类生长的必要因素,与3种微藻相比,浒苔对光照的适应性更强,在苏北浅滩特殊水体条件下适宜水深为0~0.42m,实验室培养条件下其最大相对增长率为11.91%/d;中肋骨条藻、东海原甲藻和新月菱形藻生长适宜的水深分别为0~ 0.42m,0~0.28m,0.28~0.42m,与浒苔共培养时,生长速率明显低于其单独培养,生长抑制率分别为30%~ 46%,3%~44%,25%~41%,此抑制作用与对营养盐的竞争有关,且不同微藻与浒苔竞争的作用机制不同.
[1] Smith D, Horne A. Experimental measurement of resource competition between planktonic microalgae and macroalgae (seaweeds) in mesocosms simulating the San Francisco Bay-Estuary, California [J]. Hydrobiologia, 1988,159(3):259-268.
[2] 刘 青,颜 天,周名江,等.早期发育浒苔对2株常见赤潮藻的化感效应 [J]. 海洋科学进展, 2015,33(4):529-536. Liu Q, Yan T, Zhou M, et al. Allelopathic effects ofat early stage on two representative red tide algae [J]. Advances in Marine Science, 2015,33(4):529-536.
[3] Lurling M, Van G, Scheffer M. Importance of nutrient competition and allelopathic effects in suppression of the green algaby the macrophytes,and[J].Hydrobiologia, 2006,556(1):209-220.
[4] Liu X, Wang Z, Zhang X. A review of the green tides in the Yellow Sea, China [J]., 2016,119:189-196.
[5] 张鹏燕,严振伟,钟晓松,等.浒苔生长-衰亡过程中氮形态的迁移转化过程[J]. 中国环境科学, 2019,39(5):1967-1976. Zhang P, Yan Z, Zhong X, et al. Transformation and migration of nitrogen forms during the growth-decay of[J]. China Environmental Sciencece, 2019,39(5):1967-1976.
[6] 吴 婷,赵乐毅,刘浩涤,等.浒苔对营养盐影响的初步研究[J]. 海洋环境科学, 2013,32(3):347-352. Wu T, Zhao L, Liu H, et al. Preliminary study on the influence ofon nutrients [J]. Marine Environmental Science, 2013,32(3):347-352.
[7] 庞秋婷,李 凤,刘湘庆,等.围隔实验中浒苔在不同营养盐条件下的生长比较[J]. 环境科学, 2013,34(9):3398-3404. Pang Q, Li F, Liu X, et al. Compare the growth ofunder different nutrient conditions [J]. Environmental Science, 2013,34(9):3398-3404.
[8] Kang E, Kim J, Kim K, et al. Adaptations of a green tide forming() to selected salinity and nutrients conditions mimicking representative environments in the Yellow Sea [J]. Phycologia, 2016,55(2):210-218.
[9] Cui J, Zhang J, Huo Y, et al. Adaptability of free-floating green tide algae in the Yellow Sea to variable temperature and light intensity [J]. Marine Pollution Bulletin, 2015,101(2):660-666.
[10] Li H, Zhang Y, Chen J, et al. Nitrogen uptake and assimilation preferences of the main green tide algain the Yellow Sea, China [J]. Journal of Applied Phycology, 2019,31:625–635.
[11] 吴老五,韩秀荣,吴 婷,等.重金属对浒苔吸收硝酸盐的动力学特性影响 [J]. 中国环境科学, 2016,36(4):1173-1180. Wu L, Han X, Wu T, et al. Effects of heavy metals on the uptake of nitrate by[J]. China Environmental Science, 2016, 36(4):1173-1180.
[12] 邢 飞,汪亚平,高建华,等.江苏近岸海域悬沙浓度的时空分布特征 [J]. 海洋与湖沼, 2010,41(3):459-468. Xing F, Wang Y, Gao J, et al. Spatial and temporal distribution characteristics of suspended sediment concentration in coastal waters of Jiangsu Province [J]. Oceanologiaet Limnologia Sinica, 2010,41(3): 459-468.
[13] Harrison P, Waters R, Taylor F. A broad spectrum artificial sea water medium for coastal and open ocean phytoplankton [J]. Journal of Phycology, 1980,16(1):28-35.
[14] 张美如,陆勤勤,朱庙先,等.二氧化锗(GeO2)对条斑紫菜()自由丝状体(Free Filamentous)中硅藻污染影响的研究 [J]. 现代渔业信息, 2007,22(2):24-26. Zhang M, Lu Q, Zhu M, et al. Effects of germanium dioxide (GeO2) on diatom contaminations infree filamentous [J]. Modern Fisheries Information, 2007,22(2):24-26.
[15] 张建恒,霍元子,王阳阳,等.浒苔与球等鞭金藻相互抑制的实验验证[J]. 上海海洋大学学报, 2011,20(2):211-216. Zhang J, Huo Y, Wang Y, et al. Experimental verification of mutual inhibition betweenand[J]. Journal of Shanghai Ocean University, 2011,20(2):211-216.
[16] 金送笛,李永函,倪彩虹,等.菹草()对水中氮、磷的吸收及若干影响因素[J]. 生态学报, 1994,14(2):168-173. Jin S, Li Y, Ni C, et al. Uptake byof nitrogen and phosphorus from water and some affecting factors [J]. Acta Ecologica Sinica, 1994,14(2):168-173.
[17] 韩秀荣,高 嵩,侯俊妮,等.浒苔干粉末提取物对东海原甲藻和中肋骨条藻的克生作用[J]. 生态学报, 2013,33(23):7417-7429. Han X, Gao S, Hou J, et al. Allelopathic effects of extracts frompowders on the growth ofand[J]. Acta Ecologica Sinica, 2013,33(23): 7417-7429.
[18] 史华明,石晓勇.浒苔与中肋骨条藻竞争关系的初步研究[J]. 安徽农业科学, 2013,41(16):7319-7321. Shi H, Shi X. Initial study on competitive relationship betweenand[J]. Journal of Anhui Agri. Sci, 2013,41(16):7319-7321.
[19] 侯俊妮.浒苔对三种赤潮藻的克生作用初探 [D]. 青岛:中国海洋大学, 2011. Hou J. The preliminary study of the allelopathic substances of Enteromorpha on three HABs [D]. Qingdao: Ocean University of China, 2011.
[20] 王 悠,俞志明,宋秀贤,等.大型海藻与赤潮微藻以及赤潮微藻之间的相互作用研究[J]. 环境科学, 2006,27(2):274-280. Wang Y, Yu Z, Song X, et al. Effects of macroalgae on growth of 2species of bloom microalgae and interactions between these microalgae in laboratory culture [J]. Environmental Science, 2006, 27(2):274-280.
[21] 崔 峰.浒苔对赤潮微藻的化感抑制作用以及化感物质的结构鉴定 [D]. 上海:上海海洋大学, 2014. Cui F. Studies on allelopathic effects ofon red tide microalgae and allelochemicals identification [D]. Shanghai: Shanghai Ocean University, 2014.
[22] 史华明.浒苔对氮磷吸收和释放及与中肋骨条藻竞争关系的初步研究[D]. 青岛:中国海洋大学, 2009. Shi H. The preliminary study ofabsorption and release properties of nitrogen and phosphorus and the competition with[D]. Qingdao: Ocean University of China, 2009.
[23] Nan C, Zhang H, Zhao G, et al. Allelopathic interactions between the macroalgaand eight microalgal species [J]., 2004,52:259-268.
[24] 王松波,余俊爽,曹艳敏,等.光照和营养盐对浮游动物和浮游植物生物量及其营养联系的影响[J]. 生态环境学报, 2018,27(6):1122- 1127. Wang S, Yu J, Cao Y, et al. Light and nutrients as determinants of zooplankton and phytoplankton biomass and their impacts on pelagic trophic linkage [J]. Ecology and Environmental Sciences, 2018,27(6): 1122-1127.
[25] 程丽巍.三种大型海藻对海水中营养盐供应变化的生理响应研究[D].南京:南京农业大学, 2010. Cheng L. Study on the physiological responses of three marine macroalgae to the change of nutrients supplies in seawater [D]. Nanjing: Nanjing Agricultural University, 2010.
[26] Sun K, Li R, Li Y, et al. Responses ofto short-term nutrient enrichment under light and dark conditions [J]., 2015,163:56-62.
[27] Thingstad T, Krom M, Mantoura R, et al. Nature of phosphorus limitation in the ultraoligotrophic eastern Mediterranean [J]. Science, 2005,309(5737):1068-1071.
[28] 高 嵩,石晓勇,王 婷.浒苔绿潮与苏北近岸海域营养盐浓度的关系研究 [J]. 环境科学, 2012,7:2204-2209. Gao S, Shi X, Wang T. Variation of nutrient concentrations at the inshore coastal area of northern Jiangsu province and the occurrence of green tide caused by[J]. Environmental Science, 2012,7:2204-2209.
[29] 王宗灵,李瑞香,朱明远,等.半连续培养下东海原甲藻和中肋骨条藻种群生长过程与种间竞争研究[J]. 海洋科学进展, 2006,24(4): 495-503. Wang Z, Li R, Zhu M, et al. Study on the growth process and interspecific competitions of Pandunder semi-continuous culture [J]. Advances in Marine Science, 2006,24(4):495-503.
[30] 李瑞香,朱明远,王宗灵,等.东海两种赤潮生物种间竞争的围隔实验 [J]. 应用生态学报, 2003,14(7):1049-1054. Li R, Zhu M, Wang Z, et al. Mesocosm experiment on competition between two HAB species in East China Sea [J]. Chinese Journal of Applied Ecology, 2003,14(7):1049-1054.
[31] 李鸿妹,石晓勇,丁雁雁,等.光照对东海典型赤潮藻生长及硝酸还原酶活性的影响 [J]. 环境科学, 2013,34(9):3391-3397. Li H, Shi X, Ding Y, et al. Illumination's effect on the growth and nitrate reductase activity of typical red-tide algae in the East China Sea [J]. Environmental Science, 2013,34(9):3391-3397.
Competition betweenand three red tide microalgae under different light conditions.
GAO Ying-chao, YANG Xue, LIN Kun, TAN Li-ju, WANG Jiang-tao*
(College of Chemistry and Chemical Engineering, Ocean University of China, Qingdao 266100, China)., 2019,39(8):3404~3411
There are high turbidity and large nutrient content in seawater of Subei Shoal, which is considered by most scholars to be the origin of the green tide in the South Yellow Sea. The competition betweenand,andfor nutrients under different light conditions (0~4500lx) was compared and the physiological state ofand three microalgae at different depths was evaluated combining with the actual illumination of Subei Shoal. The competition betweenand microalgae is one of the key factors determining which kind of algal bloom. The results showed thatis more adaptable to light compared with the three microalgae, the maximum average relative growth rate was 11.91 %/d. When microalgae were co-cultured with, the growth was significantly inferior to that of culture alone, the inhibition rates of,andrespectively were 30%~46%, 3%~44%, 25%~41%, and the responses of different microalgae on competition were different. The nutrients monitoring data indicated that the competition for limited nutrients was the main competitive mechanism betweenand microalgae. This result provided a basis for the outbreak mechanism of the green tide in Subei Shoal.
;microalgae;light intensity;nutrient;competition
X172
A
1000-6923(2019)08-3404-08
高颖超(1994-),女,山东淄博人,硕士研究生,主要从事海洋生态化学研究.
2019-01-21
国家重点研发计划项目(2016YFC1402101);国家自然科学基金资助项目(41876078);中央高校基本科研业务费资助项目(201713059)
, 教授, jtwang@ouc.edu.cn
