锌指蛋白ZNF222在滋养层细胞迁移与侵袭中的功能
2019-07-30刘建兵赵皓琦陈西华郝建卿徐祥波王介东
刘建兵 赵皓琦 周 芳 陈西华 郝建卿 徐祥波 贺 斌* 王介东
1.山西医科大学基础医学院(太原,030001);2.国家卫生计生委科学技术研究所
人类胎盘在进化中形成了很强的侵袭能力,胎盘最重要的细胞是滋养层细胞[1],其具有的侵袭性可侵入子宫内膜将胎盘锚定在子宫,还能侵入并重塑子宫螺旋动脉,实现整个妊娠过程中从母血中汲取胎儿所需物质,保证胎儿的正常发育和生长[2],而滋养层细胞的行为异常有可能导致妊娠疾病,几乎2/3的妊娠失败都与滋养层细胞异常的侵袭行为有关[3]。研究胎盘侵袭机制,有助于对这类疾病的认识和理解。胎盘功能正常发挥既需要旧基因的表达,更需要进化产生的新基因表达[4]。人类胎盘的侵袭能力在进化中可能伴随着新基因的产生,这些基因可能在人类胎盘特质的形成中发挥重要作用。锌指蛋白是一类具有手指状结构域的转录因子,人类多数锌指蛋白基因在免疫组织或生殖系统高表达[5-7]。有研究报道,ZNF222是在狭鼻猴类保守表达的锌指蛋白基因[8],在人类有表达而在啮齿类中不表达[9]。本文对ZNF222基因在人滋养层细胞侵袭功能中的作用进行初步研究,以期为人类胎盘侵袭能力形成的分子机制提供实验依据。
1 材料与方法
1.1 标本材料与细胞系
在知情同意的情况下,人体标本来自于北京市海淀妇幼保健院的志愿者。人滋养层细胞系HTR8/SVneo[10]培养于含有10%胎牛血清、100 IU/ml青霉素、10 mg/ml链霉素的DMEM培养基中,细胞置37℃、5% CO2中培养。本研究通过了国家卫生计生委科学技术研究所伦理委员会审查。
1.2 试剂与仪器
①免疫组织化学常规试剂耗材:ZNF222(Gene ID:7673)兔多克隆抗体(Sigma,美国),免疫组化IgG/HRP检测试剂盒(北京中杉金桥),RNA提取常规试剂TRIzol(Invitrogen, 美国)。②实时荧光定量PCR常规试剂:SYBR逆转录试剂盒(DRR081A),M-MLV逆转录酶,Random primer,RNase inhibitor,Oligo(dT),SYBR实时荧光定量试剂盒(大连宝生物)。③实时定量PCR引物:人类ZNF222 5'-TCTTGCGAGTCCTTCCGAAC-3',5'-GA AGGTCACTGCCTCCTCTG-3';GAPDH 5'-ACCACAGTCCATGCCATCAC-3',5'-TCCAC CACCCTGTTGCTGTA-3'(北京赛百盛基因技术)。④Western blot常规试剂耗材:BCA蛋白分析试剂盒(Thermo Fisher Scientific),β-Actin抗体(北京康为世纪生物)蛋白对照,RIPA蛋白裂解液(Thermo Fisher Scientific),ECL发光试剂盒(北京全式金生物),HRP 标记的二抗试剂盒(北京中杉金桥)。⑤Lipo3000及Opti-MEM培养基(Invitrogen, 美国),ZNF222 siRNA及negative control(NC)由上海吉玛制药技术有限公司设计和合成。ZNF222 siRNA共设计合成了两条,分别为:ZNF222 siRNA-1:5'-GCCAAAGAGAAG GGA AUUU TT-3',5'-AAAUUCCCUUCUCUUUGGCTT-3' ZNF222 siRNA-2:5'-GGUCUC AAGAUACC ACCAUTT-3',5'-AUGGUGGUAUCUUGAGACCTT-3',对照序列NC:5'-UUCUCCGAAC GUGUCACGUTT-3',5'-ACGUGACACGUUCGGAGAATT-3'。胎牛血清、1640培养基、HBSS(南京维森特生物),基质胶(BD,美国),CCK-8试剂盒(Dojind,日本)。⑥ Step-OneTM实时PCR扩增仪(Applied Biosystems,美国),NanoDrop 2000分光光度计(Thermo Fisher Scientific),AG22331型 PCR扩增仪(Eppendorf),DYY-III型电泳仪(北京六一仪器厂),电泳槽、转印槽、厚滤纸等(Bio-Rad,美国),RNase Free枪头、EP管(Axygen,美国),PVDF膜(Merck Millipore,德国),酶标仪(Thermo Fisher,美国),Sagecreation 凝胶成像分析系统(北京赛智创业科技)。
1.3 实验室检测
1.3.1免疫组织化学检测人绒毛、蜕膜及子宫内膜组织于4%多聚甲醛固定24 h后,常规制备石蜡切片,常规脱蜡、水化,用枸橼酸缓冲液(pH 6.0)95~98℃水浴孵育20 min行抗原修复,室温下冷却,蒸馏水浸洗5 min。切片表面滴加3% H2O2,室温孵育10 min,滴加5%非免疫原性动物血清室温封阻30 min,用ZNF222抗体(阴性对照PBS缓冲液)行一抗孵育,37℃ 60 min或4℃过夜,用相应二抗试剂盒,按照说明书方法孵育二抗,二氨基联苯胺显色,适时终止。经过PBS清洗后,用苏木精复染细胞核,脱水,中性树脂封片。
1.3.2实时荧光定量PCR在6孔板中,处理好的HTR8/SVneo细胞,用 TRIzol法提取总RNA并与Random primer及Oligo d(T)混合, 70℃ 10 min,再迅速置于冰上冷却2~3 min,配制常规逆转录体系,30℃反应10 min,42℃保温60 min,70℃反应15 min后冰上冷却,得到RNA反转后的cDNA溶液。将转染前后的HTR8/SVneo细胞的cDNA文库,用实时荧光定量PCR检测ZNF222 mRNA 表达 ,GAPDH作为内参,利用ΔΔCt法统计 mRNA 相对表达量。
1.3.3Westernblot检测在长满HTR8/SVneo细胞的6孔板中,各孔加入蛋白裂解液150~200μl,加入适量蛋白酶抑制剂,置冰上30 min,每5 min前后左右晃动一次,使裂解液和细胞充分接触,转移至离心管,12 000转/min,5 min,取上清液移至新离心管。将20~30 μg蛋白质进行10%浓度SDS-PAGE胶电泳,转移至PVDF膜后,利用 Western blot方法分别进行目标蛋白ZNF222(1:300)与内参蛋白β-actin(1:2 000)抗体杂交,ECL发光试剂盒检测。
1.3.4细胞增殖、迁移及侵袭能力检测前1天在6孔板中每孔接种2×105个细胞,利用Lipofectamine 3000转染ZNF222 siRNA 至HTR8/SVneo细胞中;转染48h后,取适量细胞种于96孔板,每一组处理重复3次,按照CCK-8试剂盒说明进行细胞增殖能力检测;transwell检测细胞的迁移和侵袭能力,结晶紫染色,显微镜下观察细胞,计数、拍照。
1.4 统计学方法
2 结果
2.1ZNF222基因表达定位
ZNF222基因在人绒毛外周滋养层区域有很高表达(图1B),在人蜕膜中也有较强表达(图1C),而在人子宫内膜中表达很弱(图1D)。

A.阴性对照;B.人绒毛;C.人蜕膜;D.人子宫内膜。Bar=100 μm图1 免疫组织化学法检测ZNF222表达 (×200)
2.2 HTR8/SVneo细胞中ZNF222基因的敲降
在HTR8/SVneo细胞中转染ZNF222 siRNA后,实时荧光定量PCR法显示 ZNF222 siRNA-1、ZNF222 siRNA-2表达量明显降低(P=0.0376、0.0392)(图2); western blot法显示 ZNF222 siRNA-1、 ZNF222 siRNA-2表达量也明显减少(P=0.0423、0.0415)(图3)。
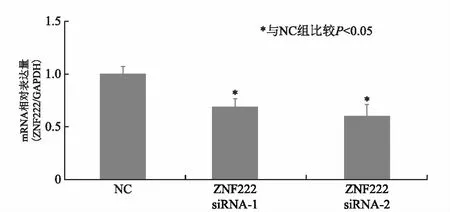
图2 RT-PCR法检测转染ZNF222 siRNA后ZNF222 mRNA表达

图3 Western blot方法检测转染ZNF222 siRNA后ZNF222蛋白水平
2.3 ZNF222对HTR8/SVneo细胞的增殖、迁移及侵袭能力的影响
ZNF222的敲降未改变细胞的增殖行为,ZNF222 siRNA-1(P=0.610)、 ZNF222 siRNA-2(P=0.277)(图4);利用transwell小室检测HTR8/SVneo细胞的迁移影响表明,ZNF222的敲降使HTR8/SVneo细胞的迁移能力减弱,ZNF222 siRNA-1(P=0.0265)、ZNF222 siRNA-2(P=0.00586);利用铺有基质胶的transwell小室检测HTR8/SVneo细胞的侵袭能力影响表明,ZNF222的敲降抑制了HTR8/SVneo细胞的侵袭能力, ZNF222 siRNA-1(P=0.0327)、 ZNF222 siRNA-2(P=0.0412)(图5)。
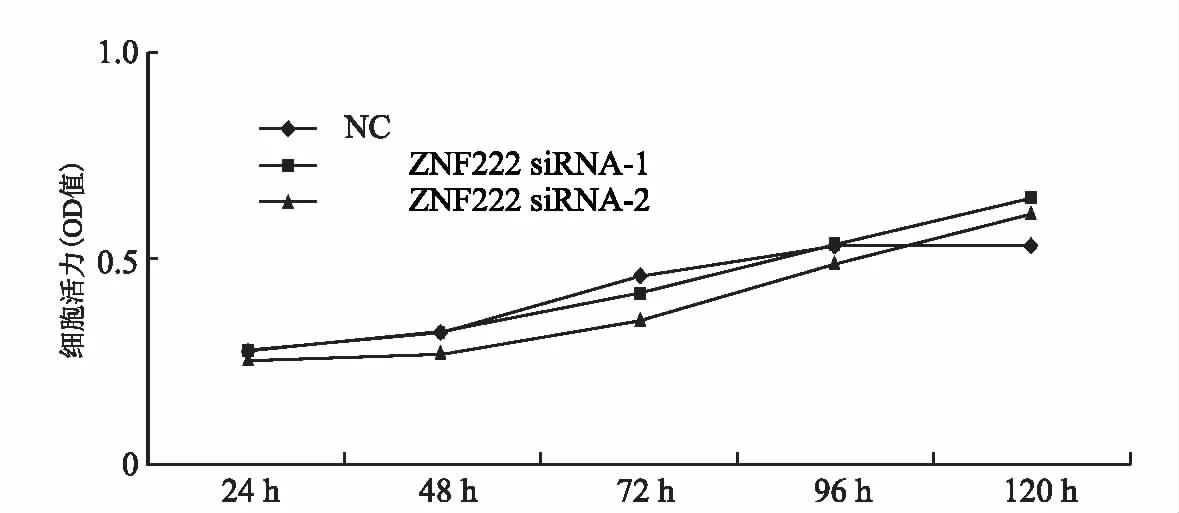
图4 敲降ZNF222后不同时间CCK-8法检测HTR8/SVneo细胞生长

图5 ZNF222的敲降对HTR8/SVneo细胞迁移和侵袭能力的影响
3 讨论
锌指蛋白是一大基因家族,分为C2H2、C4、C6、 C4HC3、C3HC4、C2HC、C3H以及联合型等多种类型,其中C2H2型最为多见,含该结构蛋白占人类蛋白总数的2%、700个左右[11],在DNA、蛋白质和RNA的相互识别中发挥重要作用。ZNF222属C2H2型锌指蛋白,位于人类19q13.31,全长7787bp,是一个编码蛋白的基因。Shannon等[8]研究表明,ZNF222与ZNF221,ZNF155、ZNF230、ZNF223、ZNF284、ZNF224、ZNF225、ZNF234及ZNF226聚集于同一段染色体中,其中人的ZNF226及ZNF234与小鼠的Zfp61相似度最高,78%~81%的氨基酸序列完全一致,而ZNF222、ZNF221,ZNF155、ZNF230、ZNF223、ZNF284、ZNF224、ZNF225在小鼠中没有极其相似的基因,但与Zfp61有一定相似[9]。由此推测,人的这8个ZNF基因可能是由Zfp61、ZNF234及ZNF226的共同祖先基因衍生出来的新基因。然而在小鼠中,该祖先基因没有衍生出新的基因或丢失了祖先基因的复制体,从而导致小鼠中只有Zfp61单个基因。
目前,有关ZNF222表达的相关报道很少。Shannon等[9]研究显示,ZNF222在人的心脏、胎盘、骨骼肌、胰腺、小肠、卵巢、脾脏、前列腺及胸腺中表达量较高,睾丸、结肠、白血球、脑及肝脏中表达量较少,在肺和肾脏中表达很少或没有表达。本研究中,应用免疫组织化学的方法显示ZNF222在人妊娠早期绒毛有很高的表达,特别是在绒毛外周有很强的表达,而在蜕膜及非妊娠状态下的子宫内膜组织中表达很弱甚至没有。这种表达模式提示ZNF222可能在人类妊娠过程中有一定作用,特别是在胎盘形成过程中滋养层细胞侵袭中可能具有重要作用。
锌指蛋白在真核生物中表达广泛, 参与细胞的分化、增殖和凋亡等多种重要生命过程。在基因表达过程中,锌指蛋白作为转录因子在DNA、蛋白质及RNA的相互识别中发挥着重要作用[12]。目前,锌指蛋白功能研究显示,锌指蛋白在人类疾病的发生发展及肿瘤细胞行为中发挥着重要作用。如,ZNF367受miR-195调控而实现抑制肿瘤细胞的增殖、迁移及侵袭[14-15];ZNF488能够激活Wnt/β-catenin信号通路而提高鼻咽癌细胞的侵袭能力[16];ZNF224通过与DEPDC1形成复合物能够诱导膀胱癌细胞的凋亡,从而在膀胱癌变过程中发挥着重要作用[17]。本研究显示,ZNF222显著抑制了HTR-8/SVneo细胞的迁移和侵袭能力,预示ZNF222的表达可能在人类胎盘形成过程中滋养层细胞侵入子宫内膜直到子宫肌层及侵入子宫螺旋动脉等行为中发挥一定作用,也可能在滋养层分化异常的相关疾病发生中发挥着一定作用。但有关ZNF222调控滋养层细胞行为的分子机制以及在疾病发生发展中可能作用,还有待进一步研究和探索。
本研究应用免疫组织化学法显示灵长类新进化产生的锌指蛋白ZNF222在人妊娠早期的绒毛中,特别是外周细胞中高表达,并且通过基因干扰技术证实ZNF222基因沉默抑制了人滋养层细胞系HTR-8/SVneo的迁移和侵袭。为锌指蛋白在人类生殖行为中的功能研究提供了新的证据,并为人滋养层细胞生物学行为的具体机制研究提供了新的线索。
