肾缺血再灌注损伤与核仁应激的相关性研究*
2019-07-25颉鸿笙乐昌昊李嘉琪李博文况晓东肖建生
颉鸿笙, 曹 源, 乐昌昊, 李嘉琪, 李博文, 况晓东, 肖建生
(南昌大学 1第一附属医院普外科, 2基础医学院病理教研室, 3第二临床医学院, 4第一临床医学院, 江西 南昌 330006)
肾脏作为高灌注器官,对缺血及缺血再灌注均十分敏感[1]。研究发现,肾缺血再灌注损伤(ische-mia-reperfusion injury,IRI)与氧化应激和内质网应激密切相关[2-3]。近年来,核仁作为一个新的并且十分重要的细胞应激中枢,在肿瘤及心血管、神经系统等疾病中发挥重要作用[4-5],但核仁应激否参与了IRI这一过程以及如何发挥作用,还未见相关报道。狭义上,核仁应激指各种损害核糖体合成并使细胞产生应激反应的过程,故也称为核糖体应激[6-7];目前被广泛定义为各种诱导核仁结构和功能改变的异常情况[8]。研究表明,核仁应激与p53关系密切[9-10],而IRI常通过p53而诱导细胞凋亡[11-12]。因此,我们分别检测肾IRI的病理改变以及核仁应激相关的RNA和蛋白表达,旨在证实肾IRI中核仁应激的发生,并进一步探究核仁应激对肾皮质再灌注细胞的影响。希望本文能从核仁的视角,为防治肾IRI提供新的策略。
材 料 和 方 法
1 实验动物
48只昆明系雄性健康小鼠, 体质量(20±5)g,购自南昌大学实验动物中心,许可证号为SYXK(赣)2010-0002。小鼠实验前10 h禁食,自由饮水。
2 肾脏IRI模型的建立
将48只小鼠按照肾缺血时间不同随机分为缺血0 min(control)组、30 min、45 min和60 min 共4个组,每组12只。称重后用10%水合氯醛溶液腹腔麻醉(4 mL/kg),待角膜及痛觉反射消失,行俯卧位固定,腰背部肾区备皮消毒。分别于脊柱左右各0.5 cm、肋骨下缘0.5 cm处剪开皮肤和肌层,进入腹腔后间隙,游离肾周筋膜和脂肪组织,暴露肾门,用微创动脉夹一次性阻断肾动脉(control组只分离出肾门处肾动脉,并继续维持腹腔开放状态60 min),双侧肾动脉夹闭的时间差小于30 s,可见双肾在1 min内颜色变黑。按照分组时间取下夹子后,双肾立即由紫黑色“扩散样”恢复为鲜红则造模成功。逐层缝合关腹,保持室温28 ℃左右,恢复供食饮水。IRI组小鼠自取下双侧动脉夹开始计时(control组满足腹腔开放状态60 min后开始计时),24 h后取样进行相关检测,同组小鼠取样时间差控制在5 min内。
3 主要方法
3.1肾功能指标的检测 测量血尿素氮(blood urea nitrogen,BUN)和血清肌酐(serum creatinine,SCr)对IRI的肾组织进行功能评估:取下动脉夹24 h后收集小鼠眼球内眦静脉血于肝素化的EP管中抗凝,混匀后立即4 000 r/min离心10 min,取上清液于4 ℃冰箱保存,并在当天分别利用肌氨酸氧化酶法和二乙酰肟法测定样本中BUN和SCr浓度(试剂盒均购自南京建成生物工程研究所),其中吸光度使用SHIMADZU公司UV-1800分光光度计测定3次,取平均值。
3.2肾系数的检测 取眼球血后迅速引颈处死小鼠,经背部依层剪开、摘取双肾,用生理盐水冲洗后剥离包膜并称重计算。肾系数=双侧肾重(mg)/体重(g)
3.3病理组织学检测 左侧裸肾称重后用全自动脱水机进行常规固定、脱水、透明和包埋,制成厚约3 μm石蜡切片,再参考HE染色试剂盒(Solarbio)说明进行染色。封片后,每张切片在高倍镜下随机取24 个视野,参照Erdogan等[13]损伤评估,采用盲法做半定量分析:0分为无损伤;1分为轻微损伤(0~5%);2分为轻度损伤(5%~25%);3分为中度损伤(25%~75%);4 分为重度损伤(75%~100%)。
3.4Western blot实验 取右侧新鲜裸肾,将皮质组织剪碎并置于RIPA和PMSF(Abcam)的蛋白裂解液中冰浴匀浆,经12 000 r/min离心5 min后保存上清液。根据BCA蛋白浓度测定结果,用Loading Buffer按照每孔10 μL上样量配平并加热100 ℃,5 min变性。样品经常规SDS-PAGE、转PVDF膜和5%脱脂奶粉封闭后I抗4 ℃孵育过夜。次日行TBST洗膜、II抗孵育以及ECL显影液凝胶成像。其中内参照选用GAPDH,目的蛋白为p53(所有抗体均购自SAB)。
3.5RT-qPCR实验 取材来自小鼠右侧新鲜裸肾,将皮质组织剪碎匀浆,采用TRIzol Reagent改良法提取总RNA。 使用EasyScript First-Strand cDNA Synthesis SuperMix(Transgen)从总RNA合成第1链cDNA,并置于-20 ℃冰箱保存备用。 20 μL RT-qPCR体系(Transgen)包括:逆转录产物1.5 μL,上、下游引物(10 μmol/L)各0.4 μL,2×Tip Green qPCR SuperMix 10 μL,ddH2O 7.7 μL。每个样品做3个复孔,经StepOnePlus Real-Time PCR仪特异性扩增目的基因的cDNA,反应条件为:94 ℃ 30 s;94 ℃ 5 s, 60 ℃ 30 s,40个循环。所得结果使用2-ΔΔCt法分析数据,引物序列见表1。

表1 RT-qPCR引物序列
F: forward; R: reverse.
4 统计学处理
所有数据均采用SPSS 20.0录入与分析,所得结果均采用均数±标准差(mean±SD)表示。多组间数据比较采用方差齐性检验和单因素方差分析,两两比较采用LSD-t检验。以P<0.05为差异有统计学意义。
结 果
1 术后存活率的比较
48只小鼠术后存活43只,存活率为89.6%,其中control组全部存活,IRI组中因手术操作不当死亡者4只,无明显原因死亡者1只,均于关腹前失去生命体征。
2 肾IRI相关指标的比较
小鼠肾脏急性IRI后,肾组织结构和功能均有明显改变。进入腹膜后间隙,可见肾包膜紧张呈肿胀样,表面时有散在分布的出血点。与control组相比,各IRI组肾系数升高(P<0.01),且60 min组较其余IRI组明显增高(P<0.01),见表2。光镜下,HE染色显示control组肾小球、肾小管结构完整清晰,未见明显异常;IRI组病理改变显著,肾小球明显充血;肾小管上皮细胞水肿呈空泡样变,部分坏死脱落造成管腔堵塞、扩张,腔内可见明显的蛋白管型和颗粒管型;肾间质显著灶性炎细胞浸润伴出血,见图1。与control组相比,各IRI组Erdogan评分增加(P<0.01),见表2;损伤范围与缺血时间呈正相关(r=0.83,P<0.01)。同时,IRI组BUN和SCr均高于control组(P<0.01),并与缺血时间呈正相关(r=0.85,P<0.01)。与30 min组比较,45 min组和60 min组BUN和SCr显著增高(P<0.01);与45 min组比较,60 min组SCr明显增高(P<0.01),见表2。上述结果显示,小鼠急性肾IRI模型建立成功,肾组织结构和功能损害与缺血时间存在相关性。

Figure 1.The pathological changes of renal cortex with different ischemic time (HE staining, ×200).
图1 不同缺血时间对肾皮质组织的病理影响
表2 小鼠肾不同缺血时间再灌注对肾系数、病理损伤评分、BUN和SCr的影响
Table 2.The changes of renal coefficient, Erdogan scores, BUN and SCr in the mice with different ischemic time (Mean±SD.n=8)

GroupRenal coefficientErdogan scoresBUN (mmol/L)SCr (μmol/L)Control12.61±2.510.38±0.5010.38±1.8139.71±5.0430 min15.82±1.68##2.88±0.99##16.65±5.13##137.80±17.49##45 min17.52±1.93##3.33±0.70##25.14±4.79##∗∗248.04±28.12##∗∗60 min20.98±1.91##∗∗▲▲3.54±0.59##∗∗30.90±6.10##∗∗310.64±27.94##∗∗▲▲
##P<0.01vscontrol group;**P<0.01vs30 min group;▲▲P<0.01vs45 min group.
3 肾IRI中核仁应激作用的观察
在肾IRI模型建立成功的基础上,我们通过RT-qPCR和Western blot检测了核仁应激指标。与control组相比,IRI各组45S pre-rRNA和18S rRNA均表达下调(P<0.01),前者较后者更为显著;各IRI组之间随着缺血时间延长,45S pre-rRNA和18S rRNA的表达降低(P<0.01),见图2A。此结果提示核糖体RNA含量下降系rDNA转录受阻,IRI使核仁的核糖体生物合成功能紊乱,导致细胞核仁应激。同时,相较于control组,IRI各组p53蛋白含量显著上升(P<0.05),各IRI组间p53表达无显著差异,见图3。

Figure 2.Relative RNA expression detected by RT-qPCR. A: relative expression of rRNA in renal cortex with different ischemic time; B: relative expression of mRNA in renal cortex with different ischemic time. Mean±SD.n=8.##P<0.01vscontrol group;**P<0.01vs30 min group;▲▲P<0.01vs45 min group.
图2 RT-qPCR 检测 RNA 的相对表达量
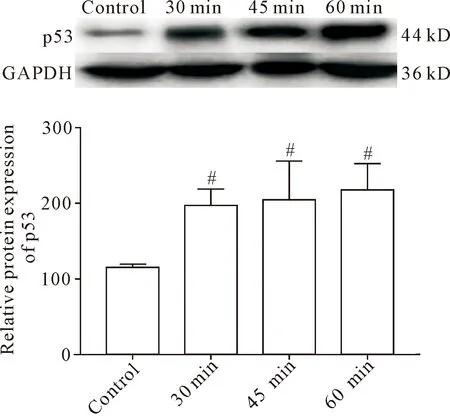
Figure 3.The protein expression of p53 detected by Western blot. Mean±SD.n=3.#P<0.05vscontrol group.
图3 Western blot 检测 p53 的蛋白表达
4 肾IRI中凋亡相关蛋白表达的比较
为了明确核仁应激在小鼠急性肾IRI中发挥保护还是损伤效应,我们利用RT-qPCR检测了细胞凋亡的指标。与control组相比,IRI各组Bax mRNA表达增加(P<0.01),Bcl2 mRNA表达下降(P<0.01);与30 min组相比,45 min组的上述改变仍具有显著差异(P<0.01),见图2B。上述结果中Bax/Bcl2比值增加,表明核仁应激在肾IRI模型中诱导肾皮质细胞凋亡。
讨 论
核仁是一种位于细胞核内的无膜亚细胞结构,主要功能是合成核糖体并响应胞内多种应激信号[14]。在真核细胞中,RNA聚合酶Ⅰ(RNA polymerase I, Pol I)和Ⅲ分别在核仁和核质中完成rDNA的转录,形成45S pre-rRNA和5S rRNA。45S pre-rRNA又在多种转录因子的参与下加工形成18S、28S和5.8S的成熟rRNA,等待与核糖体蛋白(ribosomal protein, RP)装配形成40S和60S核糖体亚基,故45S pre-rRNA及其裂解形成的下游rRNA含量共同反映Pol I的转录功能。在整个核糖体合成的过程中,大约有4 500种不同的蛋白参与其中[15],所需的物质和能量高达整个细胞的80%[16],因此细胞必须对其进行质与量的严格监管。大量研究表明[9-10]:核仁应激时RP和多种转录因子不再进行核糖体合成,而是将应激信号传递给p53,从而诱导细胞周期停滞或凋亡。例如:RPL5和RPL11会在核质中与鼠双微体2同系物(murine double minute 2 homolog, Mdm2)的中心酸性域结合,解除Mdm2对p53的泛素化抑制[17-18];核仁磷酸蛋白(nucleophosmin, NPM)也不再参与45S pre-rRNA的加工,而是因谷胱甘肽化失去与核仁内rDNA/rRNA的锚定,从而移位到核质结合Mdm2并激活p53[19]。在肾IRI模型中,抑制p53具有保护肾小管细胞的作用,提示再灌注细胞中p53与启动凋亡程序密切相关[11-12]。
本实验结果显示,与control组相比,IRI组45S pre-rRNA和18S rRNA均表达下调,反映了Pol I转录功能受阻,核仁功能障碍。同时,Western blot也检测到IRI组p53表达显著上升,证实了核仁应激的发生和信号的传递。最后,IRI组较control组Bax mRNA含量上调而Bcl2 mRNA含量下降,说明核仁应激通过p53在肾IRI中发挥损伤效应。除此之外,我们分别以Erdogan评分和BUN、SCr为衡量肾结构和功能的指标,证明了肾IRI程度与缺血时间之间存在显著相关性。
深入思考肾IRI中核仁应激发生的原因,我们认为活性氧(reactive oxygen species,ROS)的产生是一个关键因素。当创伤、休克或外科手术后氧合血流重新供应缺氧组织并导致不可逆的损伤,就会发生缺血再灌注损伤[20-21]。肾髓质和乳头细胞通过无氧酵解产生三磷酸腺苷(adenosine triphosphate, ATP),所以对低氧环境并不敏感;然而,肾皮质细胞需氧量极高,一旦肾血流中断,线粒体难以维持氧化磷酸化,胞内累积过量ROS[22-23]。在这种强烈的氧化应激状态下,不仅rDNA存在极高的损伤风险,Pol I相关的转录起始因子IA(transcription initiation factor-IA, TIF-IA)也会发生磷酸化,致使转录起始复合物组装失败[24-26]。虽然氧化应激可能导致某些定位于核仁的蛋白穿梭到核质引发核仁应激,但本文并未证明核仁应激与氧化应激直接相关,两者的因果关系有待进一步研究来阐明。
总之,我们证明了肾IRI中核仁应激的发生,希望能从保护核仁结构和功能完整的视角为减轻肾IRI损伤提供新的策略。
