水稻叶形控制基因的挖掘及分析*
2019-06-27饶玉春金哲伦胡锦春邓琳杰
饶玉春, 胡 娟, 金哲伦, 胡锦春, 邓琳杰, 周 纯, 叶 珺
(浙江师范大学 化学与生命科学学院,浙江 金华 321004)
0 引 言
粮食是人类赖以生存的物质基础,粮食的稳产、高产是社会稳定和经济发展的先决条件[1].水稻(Oryzasativa. L)是中国的主要粮食作物,自从20世纪60年代半矮杆利用和70年代杂种优势利用使我国水稻单产发生了2次革命性飞跃以后,在相当长的时间里,水稻产量的提高出现了瓶颈[2].杨守仁[3]于1996年提出了株型育种与杂种优势利用相结合的超级稻育种技术路线,近20年的实践证明,该作物育种方式进一步提升了水稻单产,已成为目前不断挖掘水稻增产潜力,突破产量瓶颈的共识策略.近年来,随着分子生物学和基因组学的不断发展,新的技术手段和方法层出不穷,与产量等优异性状相关的基因不断地被挖掘、克隆与分析,并应用于具体的育种实践中,展现出了巨大的增产潜力.
良好的个体株型是水稻高产的必备条件.水稻叶片作为光合作用的主要场所,对水稻的产量起着决定性作用.株型育种的核心思想之一就是尽可能地提高作物的叶面积指数,提高群体的光合效率和物质生产能力[4].目前,对于优良叶形性状相关的遗传和分子机理探索更是国内外株型育种研究的热点.因此,不断地挖掘与分析水稻叶形调控基因,并通过分子标记辅助选择等技术手段,将其运用到作物育种研究中,具有十分重要的学术与应用价值[5].这里主要从水稻叶长、叶宽、叶面积及叶片卷曲度等4个方面综述水稻叶形调控基因和数量性状位点(quantitative trait locus,QTLs)的研究进展,对已经克隆的叶形基因和QTLs的功能进行了梳理和总结,并对其研究前景作出了展望,以期为挖掘水稻叶形控制基因和理想株型育种提供参考,进一步提高我国水稻的产量.
1 水稻叶形概述
叶片是水稻最重要的器官之一,叶形性状关系到水稻的各项生理功能和最终产量.近年来,水稻叶形生理生化调控机制相关研究保持着较高的热度,同时也为水稻理想株型育种研究提供新的思路与参考.以下较为简略地综述了水稻叶形的分类,叶形对水稻各项生理功能的影响及水稻叶形育种研究进展与趋势,以表明叶形研究的意义与价值.
1.1 水稻叶形分类
水稻叶片形态包括叶长、叶宽、叶面积、叶周长、叶厚、叶片卷曲度、叶倾角等性状[6-10].不同叶形性状的调控基因共同参与到水稻叶片的形态结构建成和生理功能的发挥,并且不同的叶形性状在生理形态表现和遗传基础上也存在一定的关联性,如叶片窄而短的一般表现为直立叶,宽且长的则多为披垂叶,叶片卷曲有利于叶片的直立等[11-12].
1.2 叶形对水稻生理功能的影响
叶形的改变会影响到水稻的光合作用、蒸腾作用和抗逆性等多项生理功能,极大地影响到水稻的生长与产量[12].比如叶片主维管束(中脉)的发育情况直接影响到叶片的直立性和披垂度[13].叶片的挺拔度可以通过适当改良叶片宽度等性状而得到改善,进而调整整个植株的受光状态,有效增加叶面积指数,进一步挖掘出水稻的产量潜力.
1.2.1 叶形对水稻光合速率的影响
水稻叶片的光合作用受到多种叶形性状共同影响.从光合有效辐射传输角度看,叶片卷曲会减小冠层上部的遮光叶面积,进而改善中、下部冠层的受光条件.从整个冠层结构来看,叶倾角和叶片卷曲度会使冠层的叶面积密度发生改变[14].叶片的适度卷曲能够有效保证叶片的直立性,改善叶片的受光状态,增强光合,延迟叶片衰老,有效解决叶长过长导致披垂度增加的问题.直立叶片两面受光,透光率较披垂叶高,有效提升了叶面积指数,利于产量的增加.
松岛省三等[15]提出的“理想稻”株型育种中便指出上部三叶要短、厚、直立,并通过研究发现直立叶群体比弯曲群体的光合效能要多出11%~17%.郎有忠等[16]对卷叶的研究得出,卷叶的群体光合速率在适宜的高密度条件时大大超出展叶.孙虎声[17]的研究得出,卷叶的气孔导度与胞间CO2浓度均大于展叶,卷叶的光合作用速率大于展叶.角田重三郎[18]曾证明,厚叶品种中含有更多的叶肉细胞和 RUBP 羧化酶,其光合强度明显地高于薄叶品种.
1.2.2 叶形对水稻蒸腾速率的影响
叶形性状对蒸腾作用的影响相关研究相对较少.孙虎声[17]的研究表明,在齐穗前,卷叶与展叶的叶片蒸腾速率没有显著差异,但在齐穗后,卷叶的蒸腾速率一般都比展叶大.其原因可能是,齐穗后的水稻卷叶品种叶片的生理活动比展叶强,蒸腾速率相应偏高,且卷叶可能使其蒸腾作用对外界环境条件变化更敏感.通过蒸腾速率对光强的响应实验表明,当光强较小时,展叶的气孔导度和蒸腾速率均小于卷叶;当光强较大时,展叶气孔导度和蒸腾速率则超过卷叶,且随光强上升的速率也比卷叶快.
1.2.3 叶形对水稻抗逆性的影响
水稻叶形性状中,叶片厚度对抗风性有一定影响,叶片较厚,其韧性较强,因而叶片厚度适当的叶片具有比薄叶片表现为更强的抗风性,不易折断.卷叶水稻品种比展叶品种更耐高温、更耐旱[18].也有研究提出,卷叶对延缓叶片衰老进程、延长叶片功能期,尤其是中下部叶片功能期具有一定的作用.
1.3 水稻叶形育种
叶形是株形的重要组成部分,在水稻育种领域,先后有多位育种专家提出了水稻高产理想株型模式,无一例外都提及叶片形态的选育.Donald[19]于1968年首次提出理想株型育种的概念,随后水稻理想株型研究日趋深入.袁隆平院士提出的上部三叶(剑叶、倒二叶、倒三叶)要“长、直、窄、凹、厚”,这些叶片形态特征意味着能够产生足够的同化物,是超高产的基础[20].杨守仁[3]于1996年提出了采用株型育种与杂种优势利用相结合的超级稻育种技术路线.株型改良与杂种优势相结合的作物育种方式经过20多年的育种实践检验,已充分证明其可行性与正确性.中国的超级杂交水稻育种计划目前已进入第5期育种攻关,目标产量指标 16 t/hm2,水稻叶形改良育种正不断助力于水稻增产潜力挖掘[20].
2 水稻叶形控制基因遗传研究
水稻叶形控制基因的相关研究对于剖析水稻叶形性状的分子遗传机理具有重要的研究价值,并且通过对叶形控制基因的定位和克隆,应用分子标记辅助选择等技术手段,能够有效聚合有利基因,培育出包含叶形在内的众多优良农艺性状的水稻新品种.近年来,水稻叶形控制基因遗传研究领域已取得了较大的进展,因此,接下来较为详细地综述了当前水稻叶形控制基因遗传分析和叶形控制基因/QTLs定位研究进展情况,筛选并举例陈述一些已克隆的叶形性状控制基因及其功能,以期为将来叶形控制基因挖掘与分析和育种研究提供参考.
2.1 叶形控制基因遗传分析
已有遗传研究结果表明,水稻叶形相关性状,如叶长、叶宽、叶面积、叶卷曲度、叶倾角等都显示为典型的数量性状,且易受到生长环境的影响[21-22].夏仲炎[23]和马达鹏等[24]报道了粳稻剑叶长、叶宽和叶面积等性状是由微效多基因控制的数量性状,且具有较高的遗传力.晏月明等[25]研究了11个籼稻杂种剑叶性状的遗传特点,认为剑叶长、宽和叶面积至少受2对基因控制,其遗传力为60%~90%.曾建平[22]检测到控制水稻叶形性状相关QTLs位点44个,有7个QTLs的贡献率大于15%,只有1个QTLs的贡献率超过20%,因而所研究的性状都为典型的数量性状,由多个微效基因控制.蔡晶[26]分别以日本晴、TN1、II-32B与田间自主发现的突变体为亲本,利用F2代分离群体,分别对剑叶的长、宽性状进行了遗传分析.结果表明:在卷叶/日本晴的F2群体中,突变体的卷叶性状应由一个不完全显性基因控制;在超长剑叶/TN1的F2代群体中,超长剑叶可能是由一个不完全显性基因控制;通过超宽剑叶突变体分别与日本晴、TN1和II-32B构建的F2代群体的遗传分析表明,该超宽剑叶性状表现为连续变异,受多基因控制.
2.2 叶形控制基因及QTLs定位研究进展
迄今为止,国内外的研究者利用各种不同类型的遗传群体,已实现较多的叶形性状QTLs定位及部分基因的克隆.根据www.Gramene.org网站提供的QTLs定位信息,目前已定位到较多水稻叶形性状相关的QTLs,其中叶长112个、叶宽111个、叶面积39个、叶卷曲度39个、叶倾角27个等.
2.2.1 叶长基因及QTLs定位研究进展
水稻叶片(倒一叶、倒二叶、倒三叶)长度是受多基因控制的复杂数量性状.根据国家水稻数据中心网站 www.ricedata.cn 公布的数据,目前已成功克隆了控制水稻叶长性状相关基因16个,分别位于1,3,4,5,6,9,12号染色体上.Koumoto等[27]通过研究分蘖芽生长受到抑制的水稻单孽突变体moc2,发现这是由于Tos17逆转座子插入到编码胞质果糖-1,6-二磷酸酶(FBP1)的第4外显子中,导致移码而提前产生终止信号.moc2突变体是FBP1功能缺失突变体.在蔗糖合成途经中,胞质FBP酶活性缺失会导致蔗糖供应不足,很可能因此导致moc2突变体分孽芽生长受到抑制,进一步导致植株矮化,叶色淡绿,叶片短而窄,穗子小、穗粒数减少等性状.Fang等[28]从EMS处理的粳稻品种中花11群体中分离得到了一个水稻茎部发育提前终止的突变体,命名为mini1.利用分离群体分组分析和图位克隆方法,Mini1基因最终被精细定位在9号染色体上48.6 kb的区间内.序列分析表明在这个区域内有一个碱基G突变为A,导致表达的氨基酸由甘氨酸(Gly)突变为天冬氨酸(Asp).与野生型相比,mini1的茎尖分生组织(SAM)分裂停止,并导致地上部生长停止,而根部没有明显变化,叶片和叶鞘变短,叶尖枯萎,突变体在灌浆后一段时间停止生长并死亡.已克隆的Mini1基因与MOC2基因在分子作用机理上存在共通之处,都是通过抑制分蘖芽或茎尖生长,导致地上部生长停止,都作为控制叶长和叶宽的微效基因,参与到叶片大小形态建成过程中(见表1).
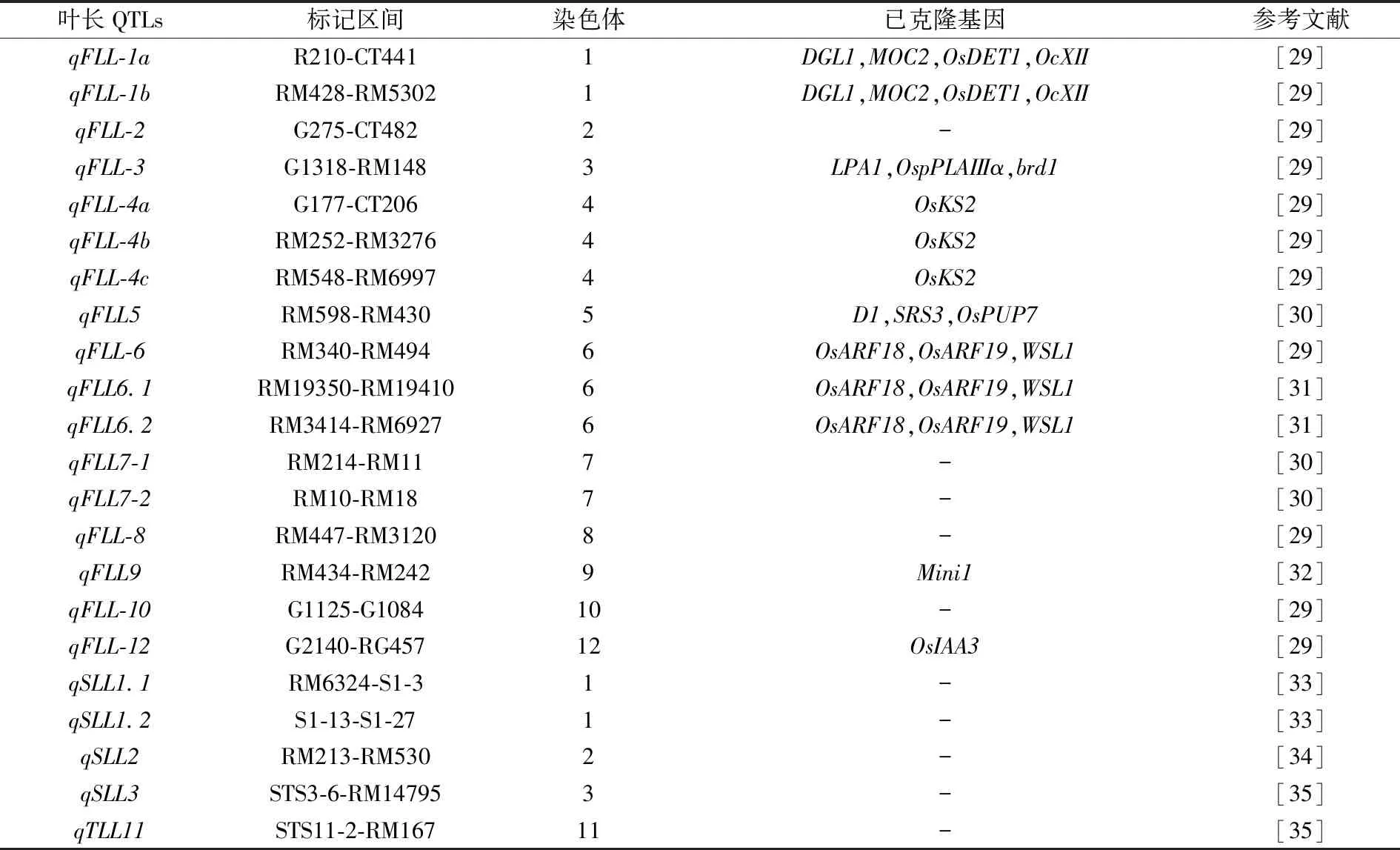
表1 叶长QTLs定位研究进展
2.2.2 叶宽基因及QTLs定位研究进展
水稻叶片(倒一叶、倒二叶、倒三叶)宽度也是由多基因控制的复杂数量性状,国内外研究者通常将叶长和叶宽同时作为研究对象,因此已定位到的QTLs数目也与叶长比较接近.根据国家水稻数据中心网站 www.ricedata.cn 公布的数据,目前已成功克隆了控制水稻叶宽性状相关基因28个,12条染色体上均有分布.Chen等[36]利用粳稻品种D50(窄叶)和籼稻品种HB277(宽叶)构建重组自交系,共鉴定到5个控制剑叶宽的QTLs.并将主效QTLsqFLW4精细定位在 RM17 483-RM17486 之间74.8 kb的区间内,该区间包含窄叶基因NAL1.nal1突变体相比野生型而言,叶片纵脉数目减少,叶片变窄,叶片长度则没有差别,而倒3,4,5节间变短,倒1,2节间长度没有差异,整体植株矮小.Li等[37]从T-DNA插入的转基因水稻中分离出一株窄叶突变体nal9.突变体在整个生长时期都表现出窄叶、苗期叶片浅绿色、株高矮化、穗变小及分蘖增加等表型.遗传分析表明,该突变体表型受单个隐性基因控制,并通过潮霉素抗性实验表明突变并不是由于T-DNA插入引起.利用图位克隆的方法分离到了NAL9基因,将NAL9基因初步定位在3号染色体上靠近分子标记RM156的位置.同时,基于籼稻和粳稻序列差异,设计RM156附近新的INDEL分子标记.通过染色体步移的方法,最终将NAL9基因定位在BAC片段OJ1212_C05上的分子标记V239B和V239G之间69.3 kb的范围内(见表2).
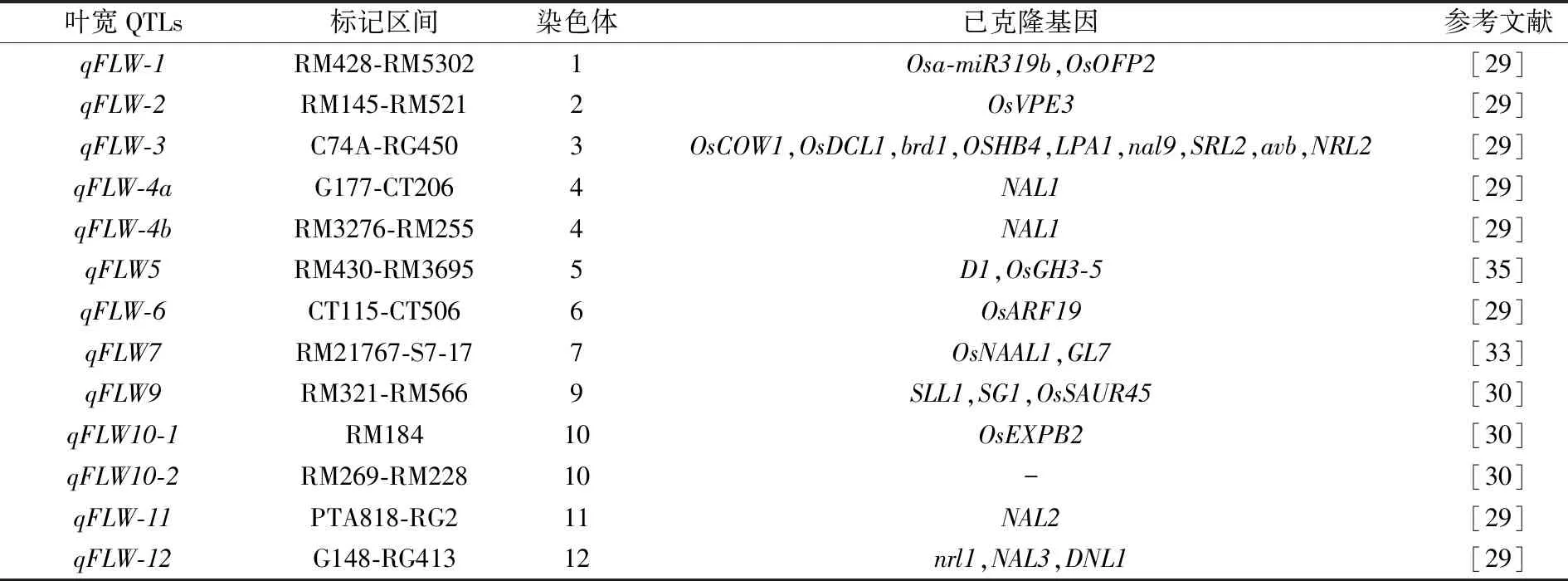
表2 叶宽QTLs定位研究进展
2.2.3 叶面积基因及QTLs定位研究进展
水稻叶面积指数的大小直接影响到水稻的光合作用、蒸腾作用等各项生理活动和最终产量,是理想株型育种的重要指标之一.叶片大小包括了叶长、叶宽、叶厚、叶周长和叶面积等性状[38],但基因具有多效型,大部分调控叶长、叶宽的基因也影响到叶面积的大小.目前,已在除11号染色体以外的其他染色体上定位到了较多的叶面积QTLs,但有关叶面积调控基因的克隆却鲜有报道.根据国家水稻信息中心网站公布的数据,只有2号染色体上的KRP1基因已被克隆.过量表达KRP1基因的转基因植株表现出叶面积稍微变小,叶片稍微变短,叶表面细胞数减少但细胞变大,饱满种子少,种子产量降低等性状.表型分析表明,KRP1过表达的植株其发育的叶片中细胞分裂减少,但被细胞增大得以部分补偿[39],具体见表3.
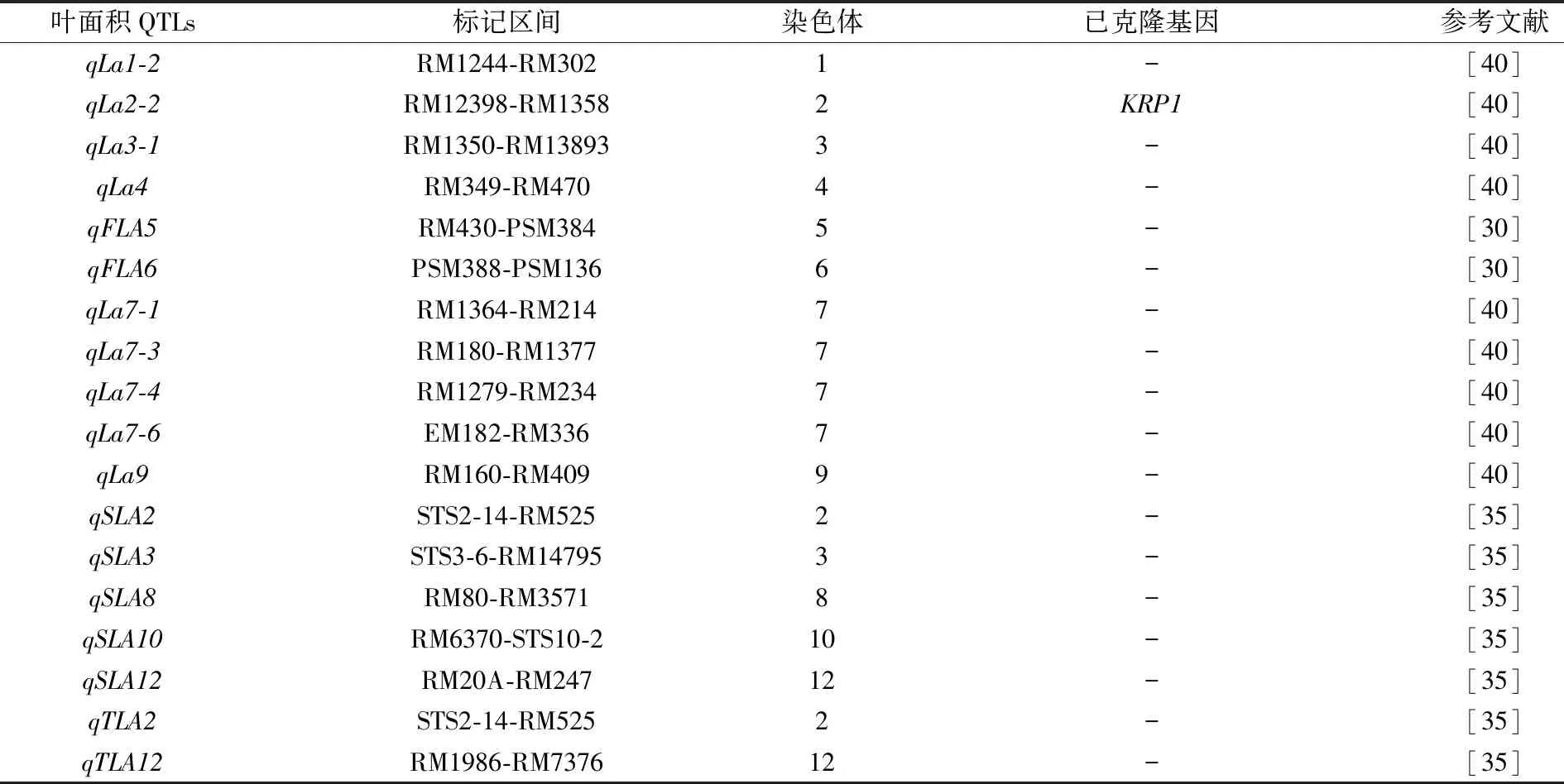
表3 叶面积QTLs定位研究进展
2.2.4 叶卷曲度基因及QTLs定位研究进展
水稻叶片的卷曲可分为内卷和外卷.卷叶的形成需要经过叶片模式建成、叶片极性建立及细胞特化等环节[41].卷叶发生的机制主要有2种:特化细胞的变异导致卷叶或者叶片泡状细胞的改变引起叶片卷曲[38].例如,Zhang等[42]报道的SLL1突变体部分出现泡状细胞异常,并且SLL1基因可通过调控叶片远轴面厚壁组织细胞的程序性死亡从而调控叶片卷曲.又如CFL1基因在叶片特化表皮细胞处过表达,可使叶片角质层受损从而导致卷叶[43].
目前已克隆的水稻卷叶相关基因已有23个,除5号染色体以外,其他染色体上均有克隆到的基因(www.ricedata.cn).徐静[44]筛选获得一株水稻卷叶突变体erl1,从苗期到成熟期都稳定表现出叶色加深、叶片极度内卷,株型矮小等性状.通过与籼稻品种TN1,ZF802,NJ06配制杂交组合,F1叶片全部为半卷叶,F2呈现“极卷叶∶半卷叶∶平展叶=1∶2∶1”的分离比,因而推断该卷叶性状为半显性单基因控制的质量性状.借助于图位克隆的手段,将ERL1基因定位于水稻第12染色体上51 kb的物理区间内.Hibara等[45]通过分离水稻adl1内卷叶突变体,发现ADL1基因编码一个半胱氨酸蛋白酶,通过促进恰当的表皮发育,在叶片和胚胎模式形成中发挥重要作用,并在整个胚胎、维管系统、叶原基、叶边缘和SAM中等组织中都有较强表达,具体见表4.
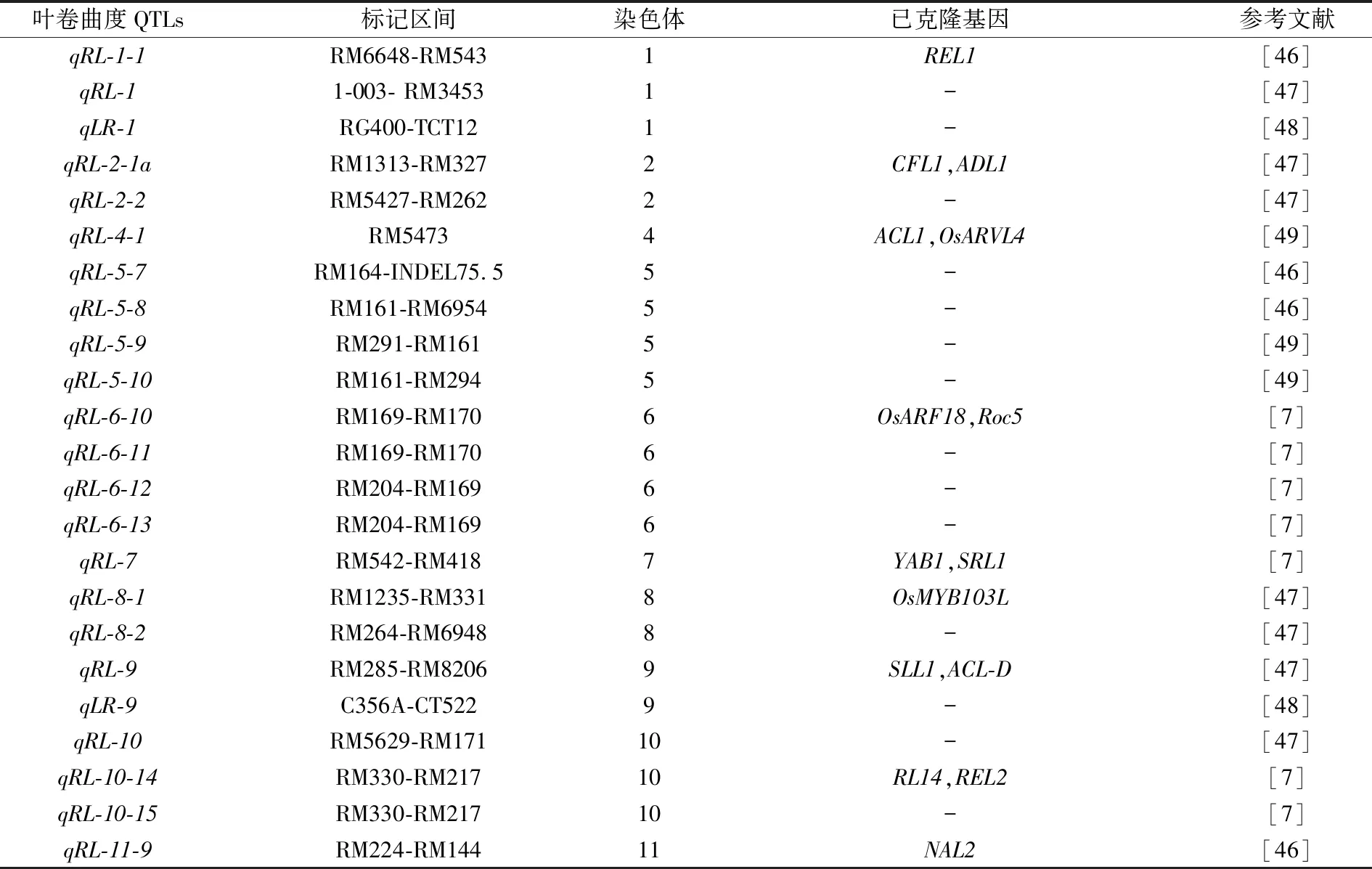
表4 叶卷曲度QTLs定位研究进展
3 总结与展望
水稻是我国主要的粮食作物,随着经济社会的不断发展,人口、环境压力与日俱增,为了保障粮食安全,确保我国水稻的稳产、高产,唯有提高水稻的单产.叶片是水稻最主要的光合器官,且与蒸腾作用和抗逆性等生理功能密切相关,是水稻产量提升的关键要素.
3.1 水稻叶形基因在育种中的应用
从水稻叶形及其调控基因角度入手改良水稻的叶形、株型,利用分子标记聚合水稻优质叶形、生理活性相关的有利基因,结合目前日益发展的分子生物学研究手段,精准设计和改良水稻新品种,将株型改良与杂种优势相结合,培育出新的性状优异、产量提升明显的水稻新品种,应用于生产实践,这是当前最有效的促进水稻产量提升的方法之一.Sakamoto等[50]研究发现,利用对油菜素内酯(BR)不敏感或与BR信号传导有关的叶片直立的矮秆突变体,可以大大提高种植密度,提高生物产量,其生物产量可比野生型增加50%.Chen等[51]培育出的高耐盐性“海水稻”——海稻86(SR 86)通过全基因组测序与比较转录组分析,发现SR 86是一个相对古老的籼亚种,拥有42 359个独有的高影响变异,并存在大量盐诱导基因,这对于水稻耐盐基因的挖掘与分析,以及更优异的耐盐性水稻育种研究具有十分重大的研究价值.但在利用分子标记辅助选择的方法改良现有的优异亲本,将良好叶形基因导入到优异亲本的过程中,须克服导入与优异叶形控制基因连锁或者不连锁的其他不利基因的问题,这样才能够行之有效地塑造理想叶形,提高水稻单产.
3.2 水稻叶形QTLs定位研究重、难点
水稻叶形相关性状大多是由多个微效基因控制的数量性状,目前已挖掘、定位和克隆所得的主效QTLs和叶形控制基因相对较少.并且基因具有多效型,同一个基因往往影响多种叶形的相关性状,如LPA1基因是目前已克隆在3号染色体上的一个调控分蘖角度和叶角的基因,同时也参与调控水稻节间宽度与厚度,茎细胞壁的厚度,谷粒和叶片的长、宽及叶面积等性状[50].QTLs定位易受遗传背景和环境的影响[29],因此有必要在不同环境条件下挖掘不同群体的QTLs.目前对于水稻叶形控制基因和QTLs的相关研究已取得了较大程度的进展,在不同环境和群体中都能检测到的QTLs表明了它们的稳定表达,而另一些QTLs则只有在某些环境下或群体中才能被检测到.另一方面,由于所使用的遗传群体和亲本材料遗传背景的差异性,以及基因组覆盖面、标记疏密程度和具体标记类型等方面的不同,QTLs定位结果往往难以直接比较,不利于进一步开展QTLs的精细定位和基因克隆.
目前叶形各性状,各种多效基因之间组合而成的复杂调控网络的剖析及相关分子遗传机理研究相对较少.因此,叶形相关性状的QTLs定位和基因克隆研究仍需更进一步地深入挖掘与分析,借助于日益发展的分子生物学与基因组学的研究手段与方法,有望实现新的突破.
