小麦穗部性状和株高的QTL定位及T6VS·6AL易位效应分析
2019-06-14胡文静高德荣陆成彬梁秀梅石宜宗程顺和
胡文静,高德荣,,陆成彬,梁秀梅,石宜宗,程顺和,
(1.江苏里下河地区农业科学研究所/农业农村部长江中下游小麦生物学与遗传育种重点实验室,江苏扬州 225007; 2.长江大学农学院,湖北荆州 434023)
迄今,已有诸多国内外学者利用不同的群体对小麦总小穗数、千粒重、穗粒数、穗长和株高进行了QTL定位[1-6]。但由于穗部性状间关系错综复杂,都是受多基因控制的数量性状,且受环境影响较大[7-8],导致前人在一些穗部性状的研究上结果差异较大,很大程度上限制了其在遗传育种中的应用。
穗粒数是重要的产量构成因素,而总小穗数和每小穗结实粒数共同决定了穗粒数,因此,通过减少不育小穗数,并增加穗顶部和基部小穗的结实粒数,可进一步增加小麦产量。Ma等[9]利用望水白和南大2419构建的RIL群体和IF2群体对小麦穗基部不育小穗数进行了QTL定位,在13个染色体区段检测到与基部不育小穗数相关的位点。Li等[10]利用川35050和山农483构建的RIL群体检测到8个控制不育小穗数的QTL。Wu等[11]利用旱选10号和鲁麦14构建的DH群体在不同干旱胁迫条件下检测到24个控制不育小穗数的QTL。Xu等[12]利用小偃54和京411构建的RIL群体在不同的氮肥和磷肥环境下检测到控制不育小穗数的QTL。Cui等[13]利用温麦8号和烟农19、温麦8号和济麦20构建的两个RIL群体检测到与可育小穗数相关的QTL。株高与小麦产量的提高密切相关,国内外学者在小麦21条染色体上相继定位出50多个株高相关QTL[14-16],且已经命名了27个主效的矮秆基因(Rht),Rht1、Rht2、Rht3和Rht10分别被克隆[17-18],其他矮秆基因研究也取得了一定的进展[19-21]。目前育种中利用最多的矮秆基因是Rht1、Rht2、Rht8、Rht9和Rht10。
扬麦17和扬麦18是江苏里下河地区农业科学研究所育成的抗病优质小麦品种和矮秆大穗白粉病免疫小麦品种,本研究利用扬麦17和扬麦18创制的206个单株组成的F2群体为材料构建遗传连锁图谱,并结合F2和F2:3群体的表型数据进行小麦穗部性状和株高QTL定位,以期发掘新的穗部性状和株高QTL,为分子标记辅助育种提供材料和技术支撑。
1 材料与方法
1.1 材 料
1.1.1 植物材料
以普通小麦扬麦18为父本,扬麦17为母本杂交获得F1代,F1自交得到206株F2植株,F2按单株收种,获得F2:3群体。扬麦18的穗顶部、基部结实性状明显好于扬麦17。簇毛麦( 2n = 14,VV) 种质来自英国剑桥植物园。抗性供体南农P045是由南京农业大学细胞遗传研究所培育的T6VS·6AL抗白粉病易位系(含PM21)。
1.1.2 引物
所用的2 403对SSR引物可覆盖小麦全基因组,全部由农业部长江中下游小麦生物学与遗传育种重点实验室(江苏里下河地区农业科学研究所)提供,由上海捷瑞生物有限公司合成。其引物序列和退火温度通过GrainGenes数据库或Somers 等(2004)公布的遗传图谱可查。
1.2 方 法
1.2.1 田间试验和表型调查
亲本材料、F2及其F2:3家系分别于2015-2016年度、2016-2017年度种植于江苏里下河地区农业科学研究所湾头基地试验田。其中,F2群体按单株种植;F2:3家系按株行种植,2次重复;行长1.3 m,行距35 cm,每行点播30粒,栽培管理同大田生产,小麦生长季未见严重病虫害和倒伏。
2016和2017年分别于小麦灌浆后期调查主茎的穗粒数、总小穗数、不育小穗数、穗顶部结实粒数、穗基部结实粒数、穗长和株高。其中,穗顶部结实粒数为顶部第一小穗、第二小穗和第三小穗结实粒数之和;穗基部结实粒数为基部第一小穗、第二小穗和第三小穗结实粒数之和。亲本P1和P2分别调查50株;F2群体调查每个单株;F2:3群体按株行调查,每行随机选取10株,取平均值,并调查2个重复。
教学中笔者发现,学生对1平方分米的表象往往停留在边长为1分米的小正方形上,观察后感悟:原来1平方分米不一定是边长1分米的正方形,可以是相应的其他图形。这一做法帮助学生突破思维定势,发展面积守恒观念。
1.2.2 DNA提取和PCR扩增
取亲本和F2群体的苗期叶片,采用常规CTAB法提取DNA,PCR体系与程序参考Taq酶(北京天根生化科技有限公司)说明书,循环数35,延伸时间为45 s,退火温度取决于不同引物。采用8%的非变性聚丙烯酰胺凝胶电泳分离PCR扩增产物,银染检测,观察记录结果并拍照保存。F2群体苗期用能够追踪T6VS·6AL易位染色体的6V染色体短臂末端共显性标记MBH1[22]进行鉴定,其正向引物序列为5′-GCCATTATAGTCAAGAGTGCACTAGCTGT-3′,反向引物序列为5′-AGCTCCTCTCGTTCTCCAATGCT-3。
1.2.3 数据分析
用 Microsoft Excel 2013对数据进行整理,用SPSS 22.0对亲本的穗部相关性状和株高进行独立样本t测验,并同时对 F2和 F2:3群体中所调查的性状进行统计和相关性分析。
1.2.4 连锁图谱的构建和QTL分析
利用JoinMap 4.0软件并采用Kosambi函数构建遗传图谱,用WinQTLCart 2.5软件选择复合区间作图法进行QTL定位分析[23],若LOD值>2.5,就认为该处存在一个有效的QTL,同时分析其加性效应和贡献率。以“Q+性状名称缩写+群体英文缩写+染色体名称+QTL次序”[24]对QTL进行命名。
2 结果与分析
2.1 表型性状及相关性分析
由表1可知,除总小穗数外,扬麦17与扬麦18在穗部相关性状和株高上均具有显著或极显著差异。其中,扬麦18的穗顶部结实粒数、穗基部结实粒数和穗粒数在表型上属于高值亲本,而扬麦17的总小穗数、不育小穗数和株高在表型上属于高值亲本。此外,穗长性状表现为扬麦17<扬麦18。F2:3群体中的偏度和峰度的绝对值都小于1,即满足正态分布。同时,各性状存在明显的双向超亲分离现象,表明均为多基因控制的数量性状。
相关性分析结果(表2)表明,在扬麦17/扬麦18的F2:3群体中,穗粒数与不育小穗数呈极显著负相关,而与总小穗数、穗顶部结实粒数、穗基部结实粒数以及穗长均呈极显著正相关,与株高呈显著正相关;穗顶部、基部结实粒数与不育小穗数呈极显著负相关,尤其穗基部结实粒数与不育小穗数之间呈高度负相关。以上结果表明,不育小穗数的减少,穗顶部结实粒数、穗基部结实粒数、穗长以及总小穗数的增加,均有利于穗粒数的增加。
2.2 遗传连锁图谱的构建
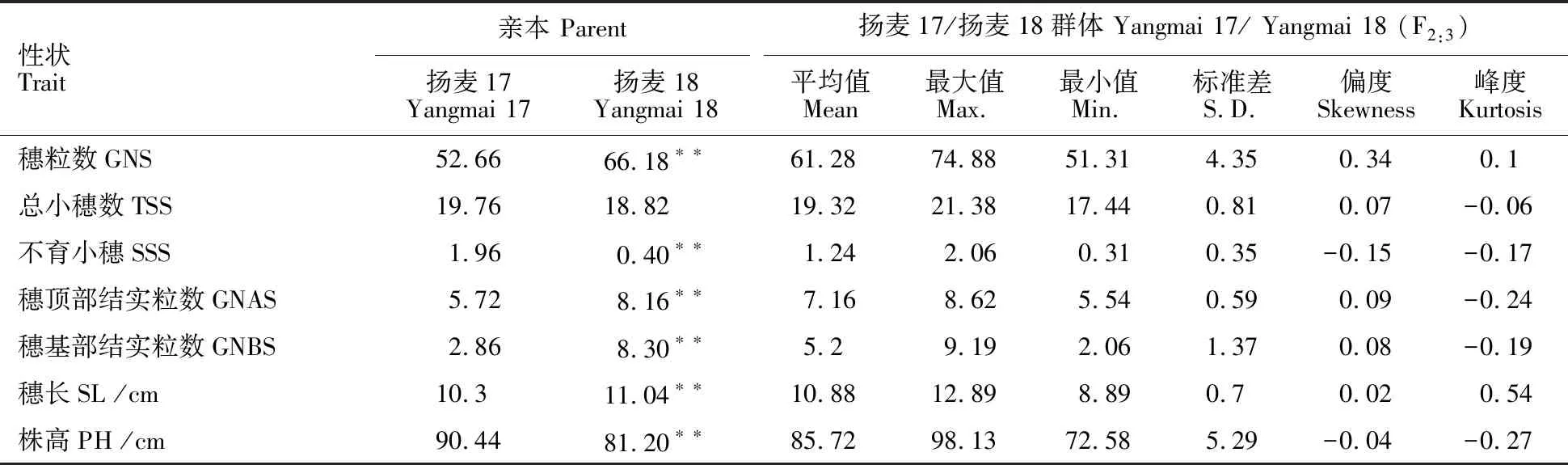
表1 扬麦17/扬麦18的F2:3群体穗部性状和株高的变异Table 1 Phenotypic variation of spike traits and plant height in the F2:3 population of Yangmai 17/Yangmai 18
GNS:Grain number per spike; TSS:Total spikelet number per spike; SSS:Sterile spikelets; GNAS:Grain number in top spikelet;GNBS:Grain number in basal spikelet; SL:Spike length; PH:Plant height.The same in tables 2-4.
** 表示与扬麦17的差异达到极显著水平(P<0.01)。
** indicates the difference reached to the extremely significant level compared with Yangmai 17(P<0.01).

表2 扬麦17/扬麦18F2:3群体中穗部各性状之间的相关系数Table 2 Correlation coefficients for all traits in the F2:3 population of Yangmai 17/Yangmai 18
*和** 分别表示相关性达到显著(P<0.05)和极显著(P<0.01)水平。
*and ** indicate the correlation reaching the significant level at 0.05 and 0.01 levels,respectively.
2.3 QTL定位分析
基于复合区间作图法,共定位到15个控制穗部性状和株高的QTL,分布在2B、2D、4B、5A、5B、7A染色体上(图1和表3)。

图1 (扬麦17/扬麦18)F2和F2:3群体穗部性状和株高QTL在染色体上的分布
2.3.1 穗部性状QTL
1个控制穗粒数的QTL位于5B染色体上的Xgpw4483~Xgpw4478标记之间,仅能在F2代检测到,可解释9.92%的表型变异,其增效等位基因来自扬麦18。3个控制总小穗数的QTL分别位于2B和5B染色体上,仅在F2:3代被检测到。其中,QTss-YY-5B位于Xgdm3~Xgpw7180之间,表型变异解释率为7.28%,增效基因来源于扬麦17;QTss-YY-2B.1和QTss-YY-2B.2分别位于Xgwm429~Xmag3635和Xgwm148~Xgpw4347之间,两个QTL的峰值在20 cM以内,推测是相同QTL,表型变异解释率分别为6.77%和9.21%,增效等位基因均来自扬麦18。2个控制不育小穗数的QTL均位于2B染色体上,仅在F2:3代检测到,分别位于Xgwm120~Xgpw4112和Xgpw4112~Xgwm501之间,Xgpw4112是二者共同紧密连锁的分子标记,两个QTL的峰值在20 cM以内,推测是相同QTL,QTL表型变异解释率分别为7.05%和7.65%,增效等位基因均来自扬麦18。1个控制穗基部结实粒数的QTL位于5B染色体上,位于Xgpw4483~Xgpw4478之间,仅在F2代检测到,表型变异解释率为6.43%,增效等位基因来源于扬麦18。未能检测到控制穗顶部结实粒数及其构成性状的QTL。

表3 扬麦17/扬麦18 F2和F2:3群体穗部性状以及株高的QTL定位结果Table 3 QTLs for spike traits and plant height in the F2 and F2:3 populations of Yangmai 17/Yangmai 18
加性效应值中,正值表示增效基因来自扬麦17,负值表示增效基因来自扬麦18。
Positive additive effect values indicate increased effects from Yangmai 17,and negative additive effect values indicate increased effects from Yangmai 18.
5个与穗长相关的QTL分别位于2B、2D、5A和7A(2)染色体上。其中,QSl-YY-7A.1、QSl-YY-7A.2分别位于Xpsp3050~Xgpw7386和Xgpw2119~Xmag794之间,仅在F2代检测到,两个QTL的峰值在20 cM以内,推测是相同QTL,表型变异解释率分别为7.39%和6.32%,增效等位基因均来自扬麦17;QSl-YY-2B位于Xgwm429~Xmag3635之间,仅在F2:3代下检测到,表型变异解释率为5.98%,增效等位基因来源于扬麦18;QSl-YY-2D和QSl-YY-5A在两代中均能被检测到,前者在两代中都位于Xcfd53~Xgwm261之间,表型变异解释率分别为9.71%和10.43%,增效等位基因均来自扬麦17,后者在两世代中分别位于Xgpw7007~Xwmc327、Xwmc327~Xbarc1182之间,表型变异解释率分别为7.99%和11.84%,增效等位基因均来自扬麦18。
2.3.2 株高QTL
3个控制株高的QTL被定位在2D、4B和5A染色体上。其中,QPh-YY-2D和QPh-YY-4B分别位于Xcfd53~Xgwm261和Xbarc1096~Xgpw4335之间,在两世代中均被检出,表型变异解释率分别为1.93%和20.78%,增效等位基因均来自扬麦17;QPh-YY-5A位于Xwmc327~Xbarc1182之间,仅能在F2代中检测到,表型变异解释率为3.84%,增效等位基因来自扬麦18。
2.4 小麦-簇毛麦易位染色体T6VS·6AL在F2群体中的传递及对小麦穗部性状的影响
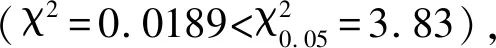

表4 两世代中含有纯合T6VS·6AL、杂合T6VS·6AL和不含T6VS·6AL的个体间穗部性状比较Table 4 Comparison of spike traits among homologous T6VS·6AL,heterozygous T6VS·6AL and non T6VS·6AL subgroups in two generations
*表示两世代中含有纯合T6VS·6AL与不含T6VS·6AL类群间的差异达到显著水平(P<0.05)。
* indicates the difference between homologous T6VS·6AL and non T6VS·6AL subgroups in two generations reaching a significant level(P<0.05).
3 讨 论
3.1 QTL位点的一致性
本研究结果表明,穗长QTL分别位于2D、5A、7A和2B上, 增效等位基因QSl-YY-2D位于Xcfd53和Xgwm261之间,这与Heidari等[25]和Sourdille等[1]的研究结果一致;增效等位基因QSl-YY-5A位于Xbarc327~Xbarc1182之间,与Liu等[26]检测到与穗长相关的QTLQSL-5A.2所属区段相同,推测是同一QTL;在7A上检测到的穗长QTL与前人报道[26-27]相距较远,推测是一个新的QTL;QSl-YY-2B所属标记区间Xgwm429~Xmag3635位于2BS上,标记Xgwm429落在侯立江[28]定位到的Qsl2B区段内,但是因为另一标记不在此区段,因此无法推断是否是同一QTL。
株高QTL分别位于2D、4B和5A上,QPh-YY-2D位于Xcfd53~Xgwm261区段内,增效等位基因来自扬麦17,已有报道显示Xgwm261可以作为2DS上矮秆基因Rht8的特异分子标记[29],推测此QTL是Rht8。QPh-YY-4B位于Xbarc1096~Xgpw4335区段内,F2:3代表型贡献率达到20.78%,增效基因来自扬麦17,经过与中国春(CS)V1.0 版本参考基因组(http://plants.ensembl.org/Triticum_aestivum/Info/Index)比对发现该QTL位于4BL上,与已报道4BS上矮秆基因Rht-B1b的物理位置相距较远,推测此处是一个新的矮秆QTL。QPh-YY-5A位于Xwmc327~Xbarc1182区段内,增效基因来自扬麦18,与Gao等[30]报道的5AS上株高QTL位置不同,与已知矮秆基因Rht12相距较远[31],推测是一个新的株高QTL。
总小穗数QTLQTss-YY-2B.1位于Xgwm429~Xmag3635区段,增效等位基因来自扬麦18,此QTL与梁秀梅等[32]检测到的总小穗数QTLQTSS-2B区段具有共同分子标记Xgwm429,推测是同一QTL。另一总小穗数QTLQTss-YY-5B,表型贡献率7.28%,增效基因来自扬麦17,所属区段为Xgdm3~Xgpw7180,未见前人报道。
5B长臂末端同时定位到穗粒数和穗基部结实粒数QTL,增效等位基因均来自扬麦18,标记区间为Xgpw4483~Xgpw4478,这与王健胜[33]定位到的控制穗粒数的QTLQGns.wa-5B.e3所属标记区段位置相近[27],推测是同一QTL。有关穗基部结实粒数QTL研究罕见,已知梁秀梅等[32]在3D上检测到穗基部结实粒数QTLQGnbs-YN-3D,其他未见报道,因此本研究定位到的QGnbs-YY-5B是一个新的穗粒数QTL。
3.2 QTL 位点的表型贡献率和稳定性
定位结果显示大多数穗部性状QTL的贡献率较低,只有QSl-YY-2D、QSl-YY-5A、QPh-YY-2D和QPh-YY-4B能够同时在两世代中被检测到,其他位点只能在单一世代被检测到,而QPh-YY-2D在F2:3世代表型贡献率达到20.78%,F2代表型贡献率只有4.9%。这是因为穗部性状大都属于数量遗传性状,由微效多基因控制,且受环境影响,并且供试群体F2和F2:3属于低世代临时性群体,表型调查误差大,导致大多数穗部性状在不同世代中检测到的QTL有差异。我们后续会增加重组自交系群体多环境试点以提高结果的可靠性和稳定性,寻找在不同环境下稳定表达的穗部相关性状QTL。
3.3 QTL位点的“一因多效性”或者紧密连锁
相关性状的QTL往往存在于相同或相近染色体区,表现出“一因多效”或紧密连锁效应,通过不同性状的QTL同时定位可以在一定程度上解释复杂性状及其构成要素或相关性状之间的关系,也在一定程度上证明了该位点的稳定性。本研究在2D、5A染色体同一区段发现了控制穗长和株高的QTL,在5B染色体同一区段发现控制穗粒数和穗基部结实粒数的QTL,在2B染色体相近区段发现控制总小穗数和穗长的QTL。利用这些“一因多效”或者紧密连锁的QTL,结合稳定表达的位点,通过回交和分子标记辅助选择,可同时聚合不同位点上控制多个性状的等位基因,从而创制出优异的小麦种质资源。
3.4 T6VS·6AL易位系的遗传效应
李桂萍等[34]的研究结果表明,T6VS·6AL易位染色体对后代的穗长和千粒重表现出一定的正向效应,对其他产量构成要素及产量没有表现出明显的影响。本研究供试亲本扬麦18与扬麦17杂交后获得的F2代中T6VS·6AL类群的穗顶部结实粒数显著高于非T6VS·6AL类群,而穗长、穗粒数和穗基部结实粒数在两个类群之间无显著差异,与以往的报道不完全一致,这是因为外源染色体导入小麦背景后产生的遗传效应有可能受小麦不同遗传背景、不同世代的影响。
F2代中,含有纯合T6VS·6AL类群的穗长、穗粒数、穗顶部结实粒数最高,含杂合T6VS·6AL类群的穗长、穗粒数、穗顶部结实粒数和穗基部结实粒数均高于另两个类群的平均值,不含T6VS·6AL类群的穗长、穗粒数、穗顶部结实粒数和穗基部结实粒数最低。而F2:3代中含有纯合T6VS·6AL类群的穗长、穗粒数最高,含杂合T6VS·6AL类群的穗长、穗粒数均高于另两个类群的平均值,初步推测6VS上携带有增加穗长、穗粒数的基因,且为部分显性。由于研究所用的群体为尚未稳定的分离群体,特别是F2群体缺少生物学重复鉴定,因此本研究获得的有关T6VS·6AL对产量性状的效应还需进一步利用稳定的重组自交系群体通过多年多点试验进行验证。
致谢:感谢中国农业科学院作物科学研究所夏先春研究员在 KASP 基因分型方面提供的技术支持!
